

Transforming representations of movement from body- to world-centric space
- PMID: 34912123
- PMCID: PMC10759448
- DOI: 10.1038/s41586-021-04191-x
Transforming representations of movement from body- to world-centric space
Abstract
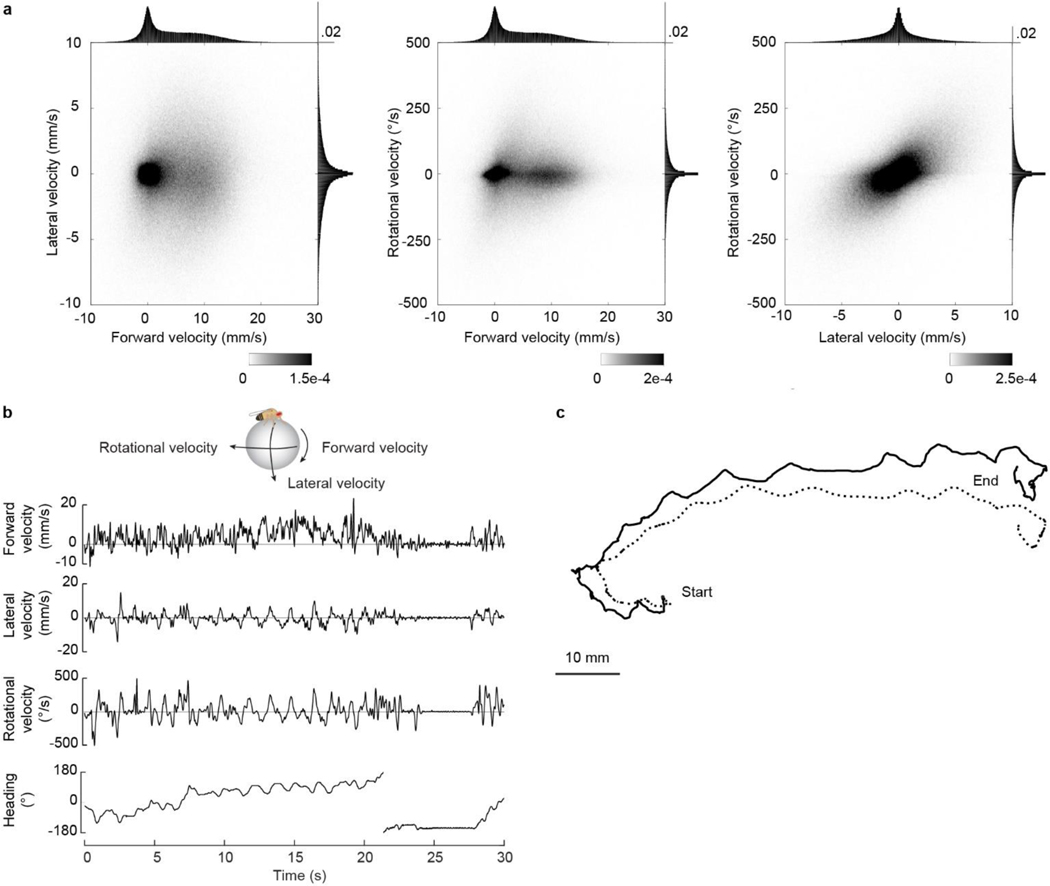
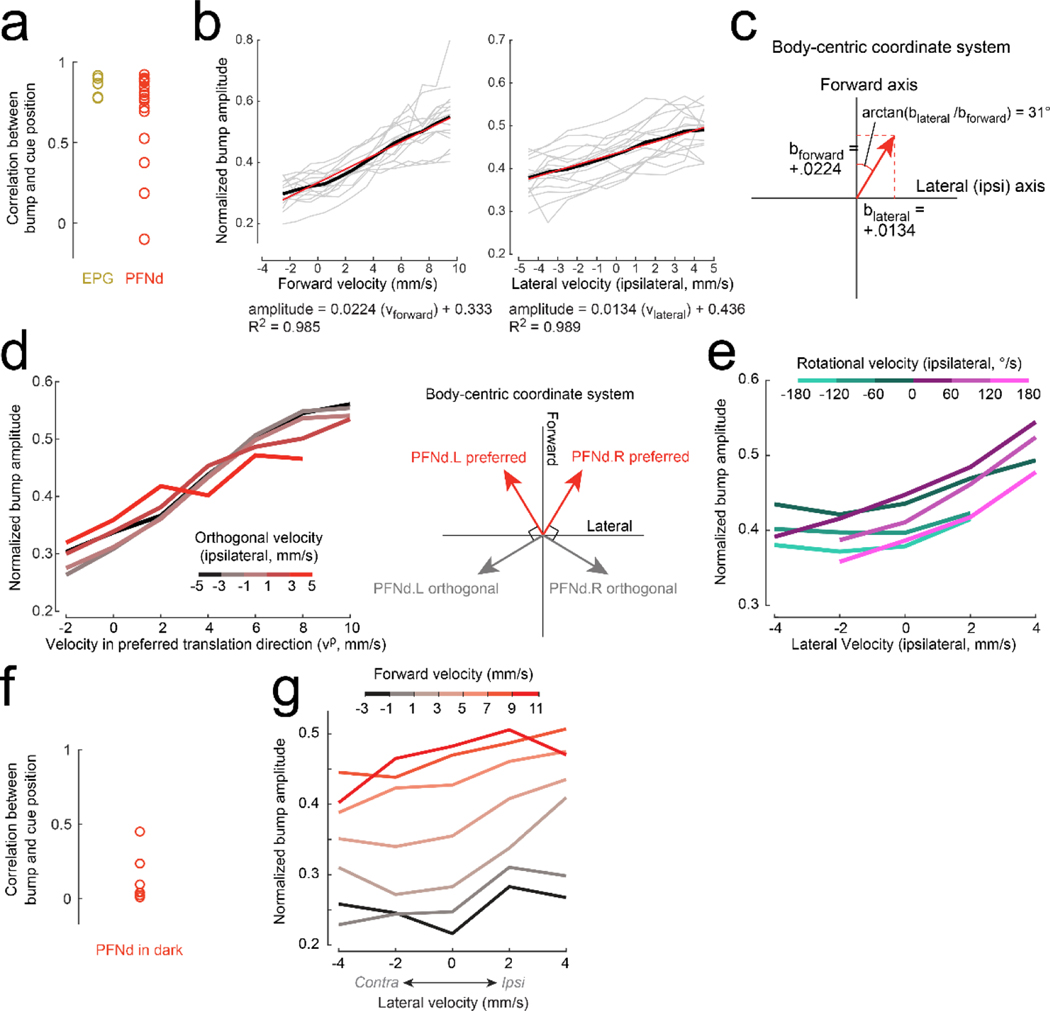
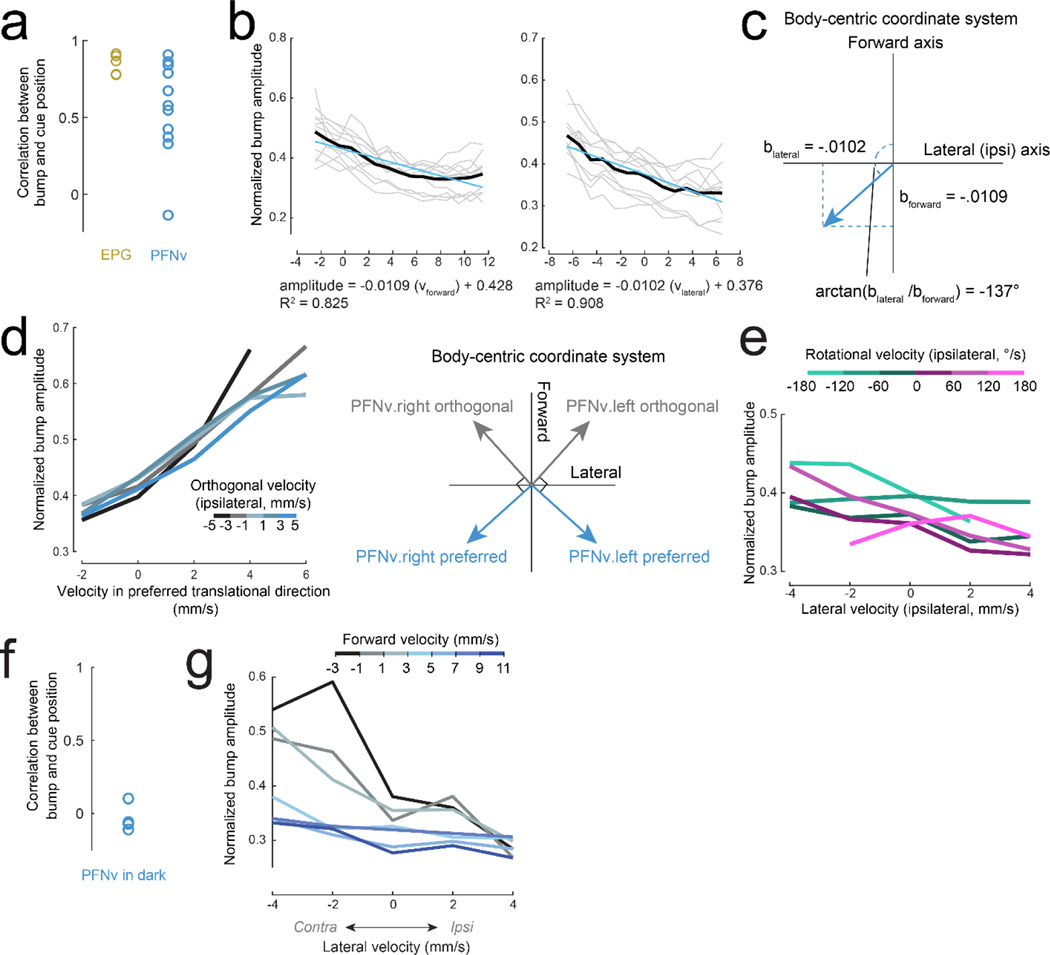
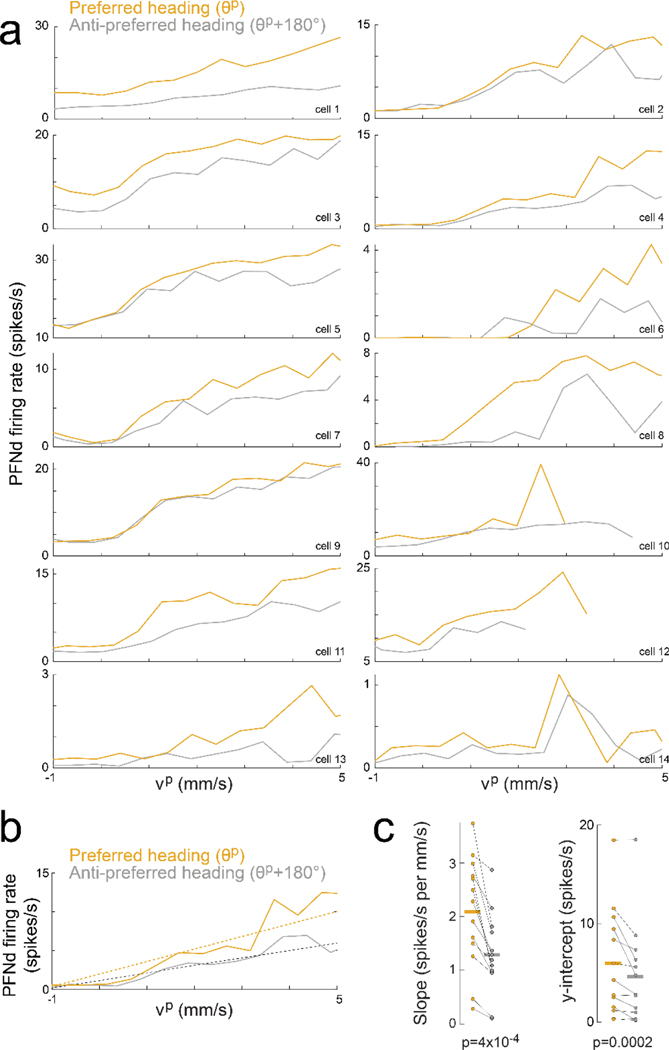
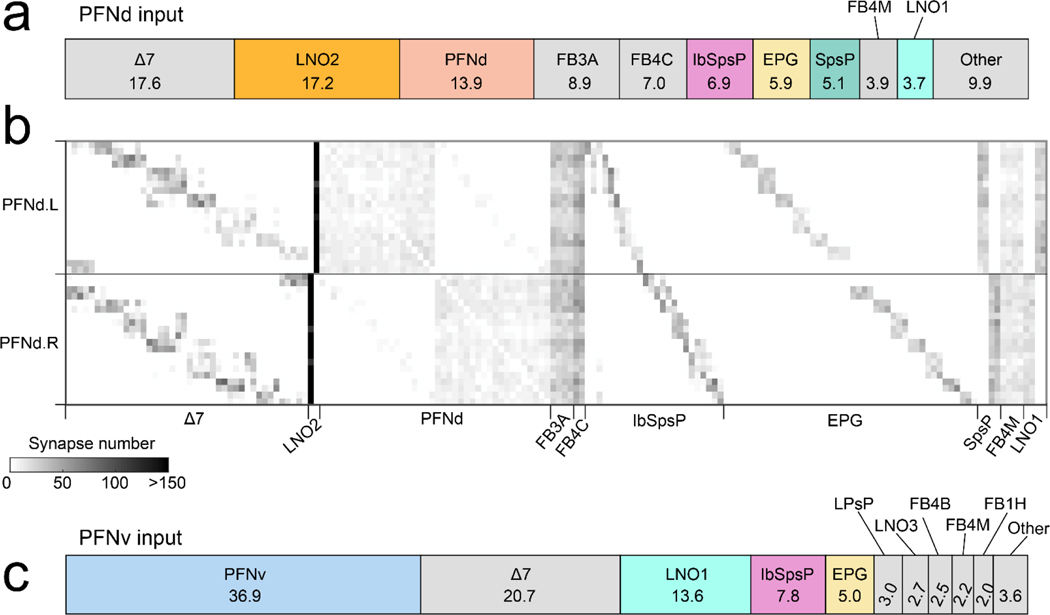
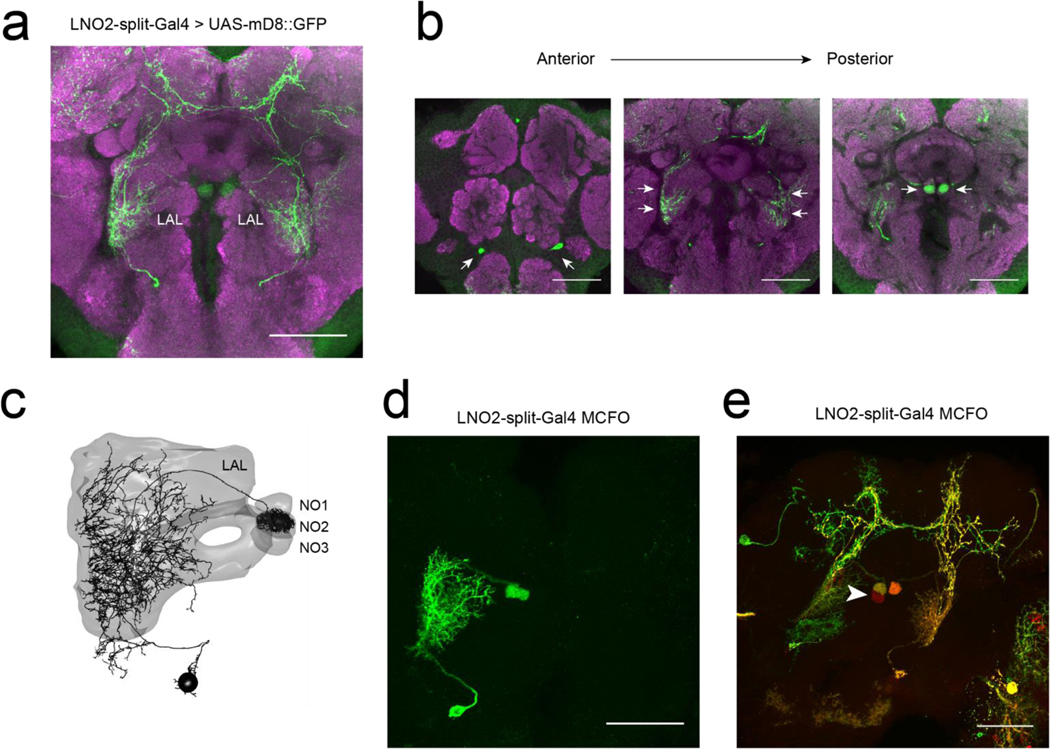
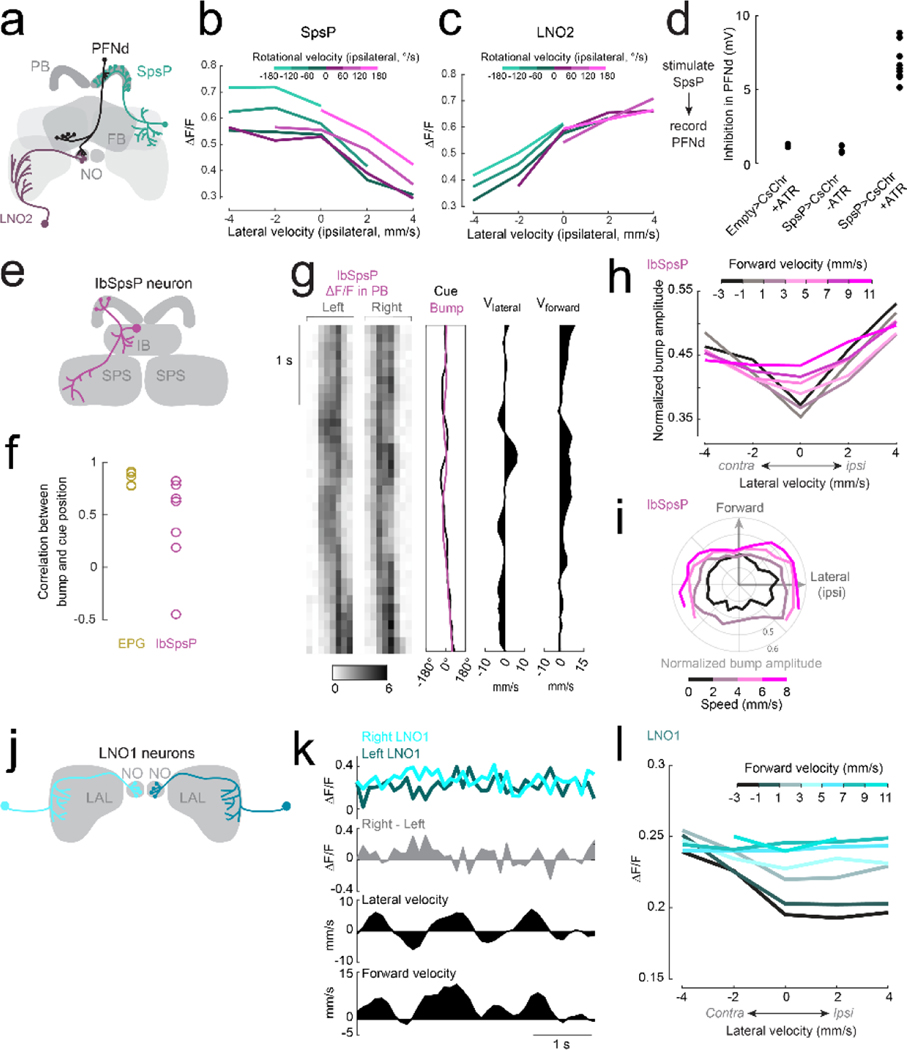
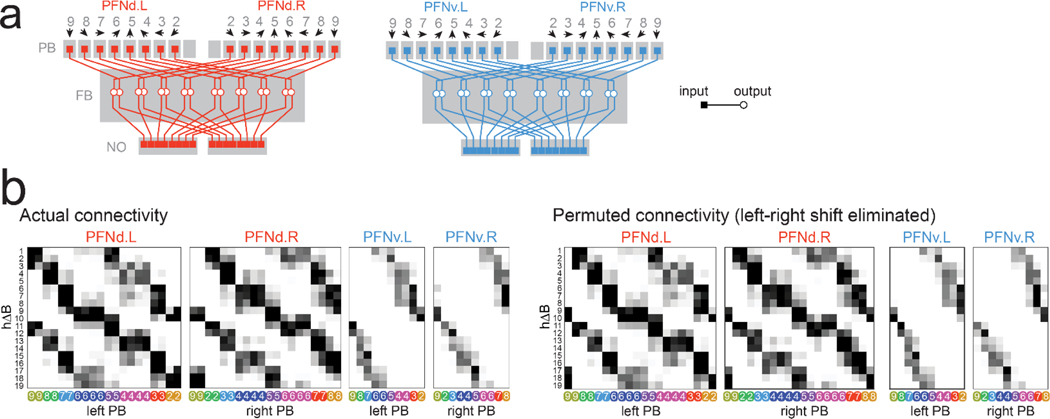
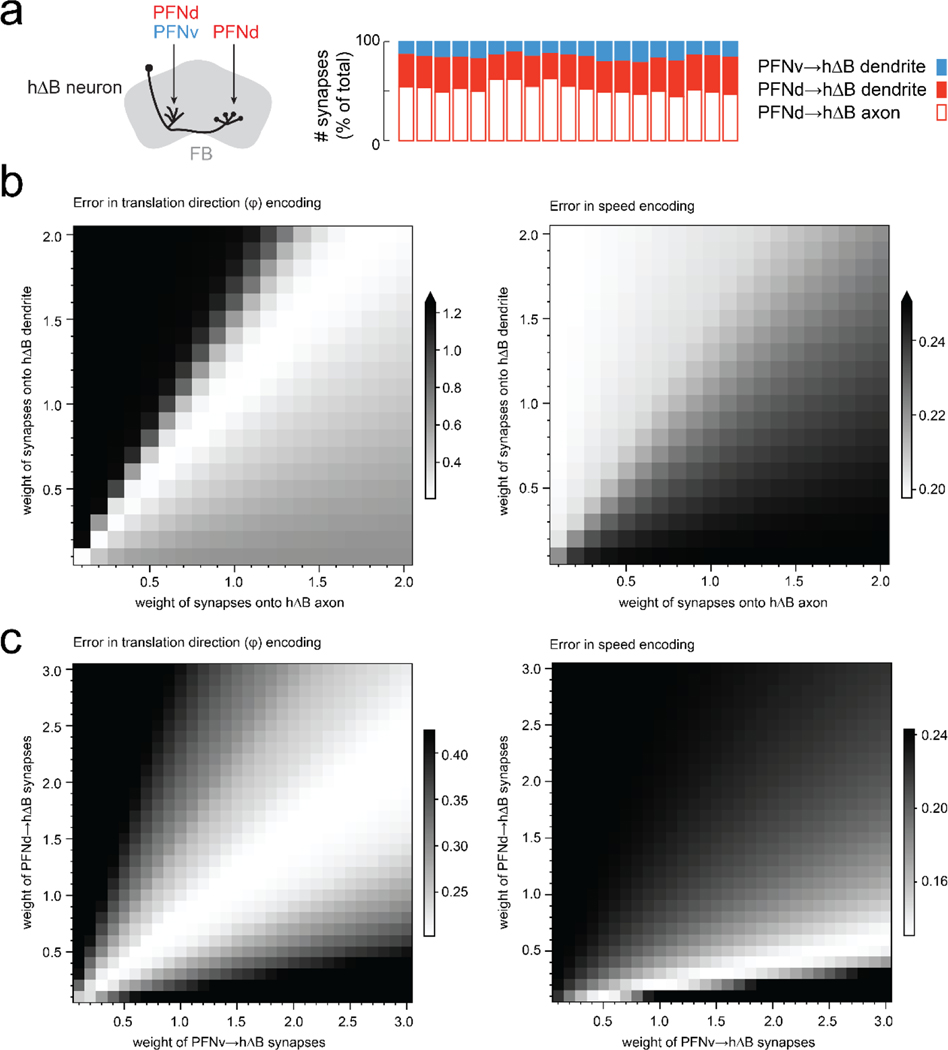
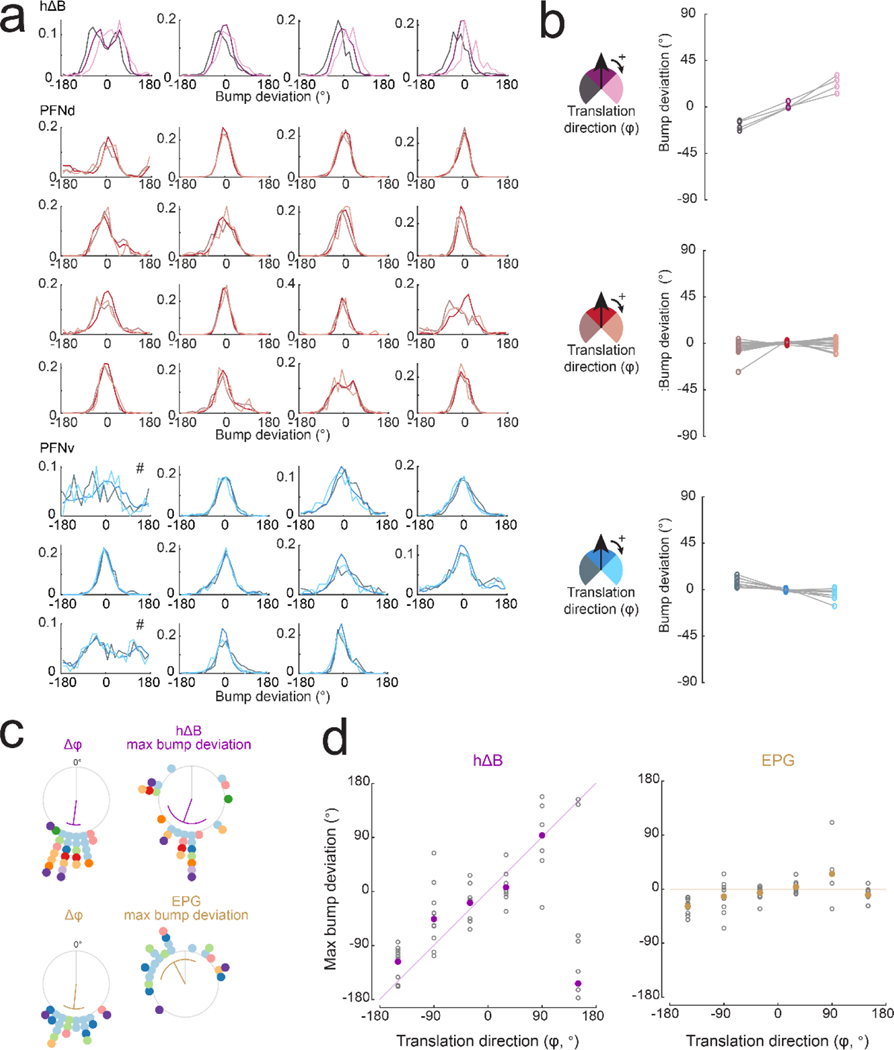
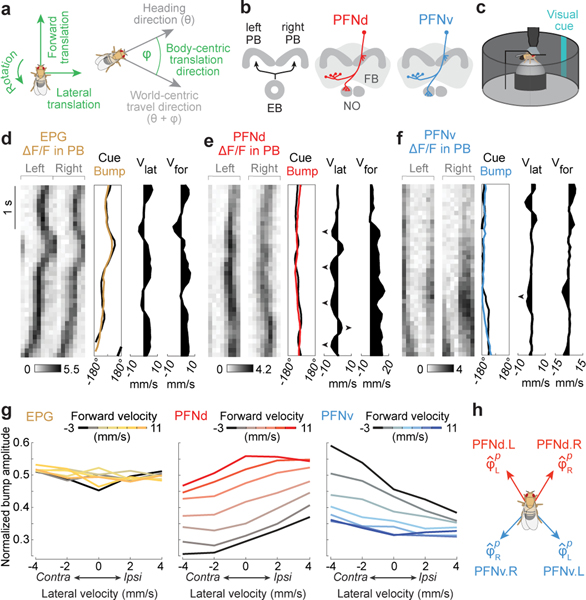
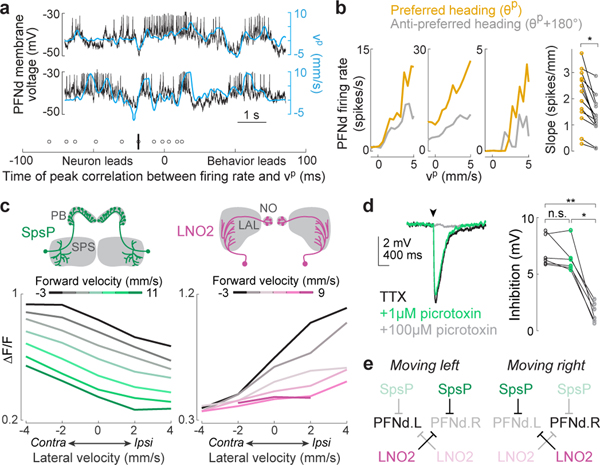
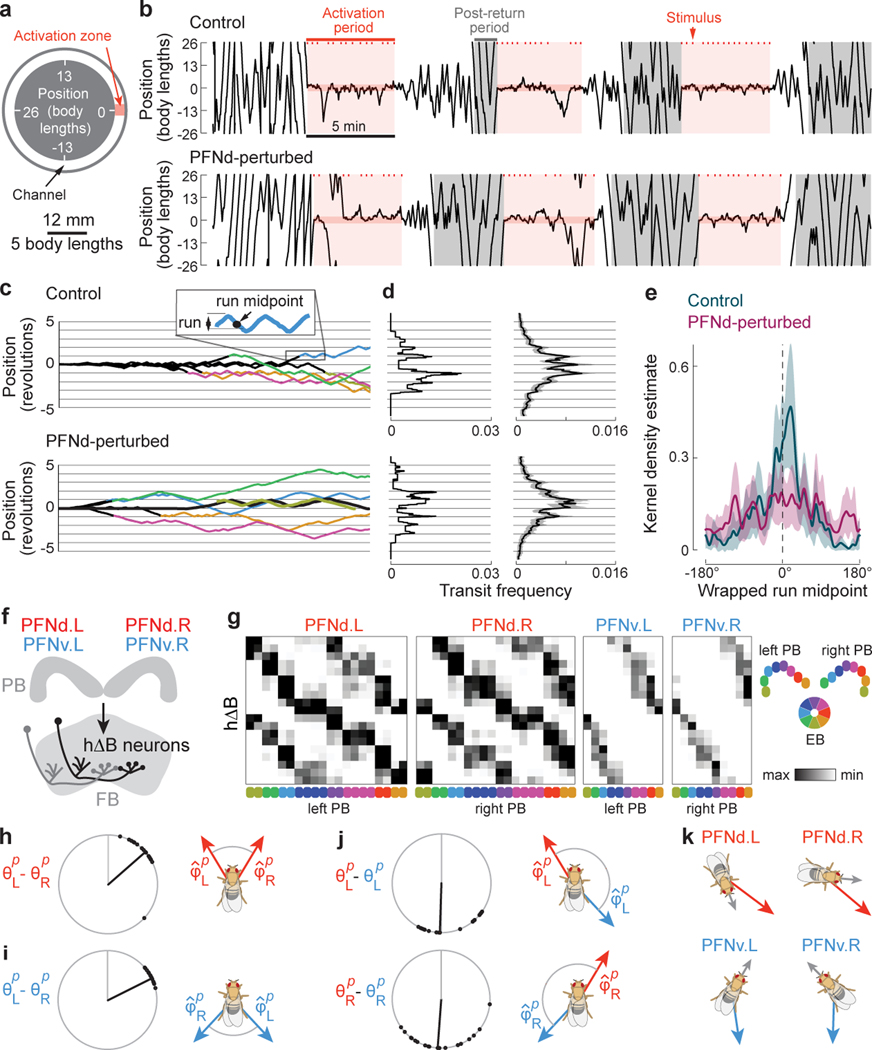
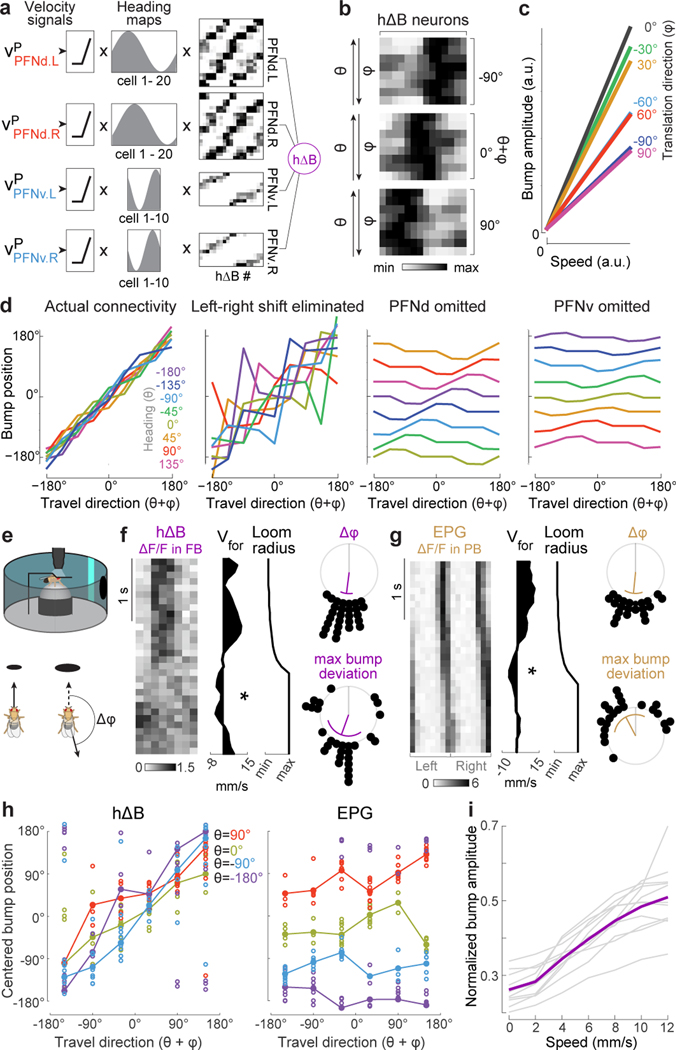
When an animal moves through the world, its brain receives a stream of information about the body's translational velocity from motor commands and sensory feedback signals. These incoming signals are referenced to the body, but ultimately, they must be transformed into world-centric coordinates for navigation1,2. Here we show that this computation occurs in the fan-shaped body in the brain of Drosophila melanogaster. We identify two cell types, PFNd and PFNv3-5, that conjunctively encode translational velocity and heading as a fly walks. In these cells, velocity signals are acquired from locomotor brain regions6 and are multiplied with heading signals from the compass system. PFNd neurons prefer forward-ipsilateral movement, whereas PFNv neurons prefer backward-contralateral movement, and perturbing PFNd neurons disrupts idiothetic path integration in walking flies7. Downstream, PFNd and PFNv neurons converge onto hΔB neurons, with a connectivity pattern that pools together heading and translation direction combinations corresponding to the same movement in world-centric space. This network motif effectively performs a rotation of the brain's representation of body-centric translational velocity according to the current heading direction. Consistent with our predictions, we observe that hΔB neurons form a representation of translational velocity in world-centric coordinates. By integrating this representation over time, it should be possible for the brain to form a working memory of the path travelled through the environment8-10.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures
References
-
- Bicanski A. & Burgess N. Neuronal vector coding in spatial cognition. Nat. Rev. Neurosci 21, 453–470 (2020). - PubMed
Additional references for Online Methods
-
- Tirian L. & Dickson BJ The VT GAL4, LexA, and split-GAL4 driver line collections for targeted expression in the Drosophila nervous system. bioRxiv 198648 (2017).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
