

ROCK2 inhibition attenuates profibrogenic immune cell function to reverse thioacetamide-induced liver fibrosis
- PMID: 34917911
- PMCID: PMC8645924
- DOI: 10.1016/j.jhepr.2021.100386
ROCK2 inhibition attenuates profibrogenic immune cell function to reverse thioacetamide-induced liver fibrosis
Abstract
Background & aims: Fibrosis, the primary cause of morbidity in chronic liver disease, is induced by pro-inflammatory cytokines, immune cell infiltrates, and tissue resident cells that drive excessive myofibroblast activation, collagen production, and tissue scarring. Rho-associated kinase 2 (ROCK2) regulates key pro-fibrotic pathways involved in both inflammatory reactions and altered extracellular matrix remodelling, implicating this pathway as a potential therapeutic target.
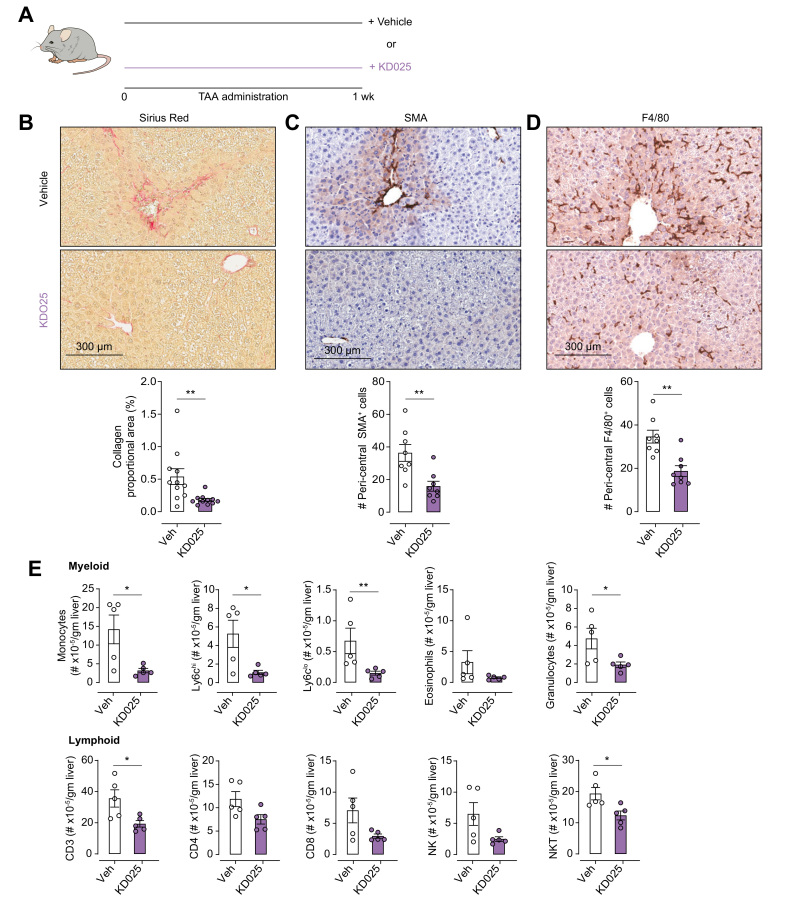
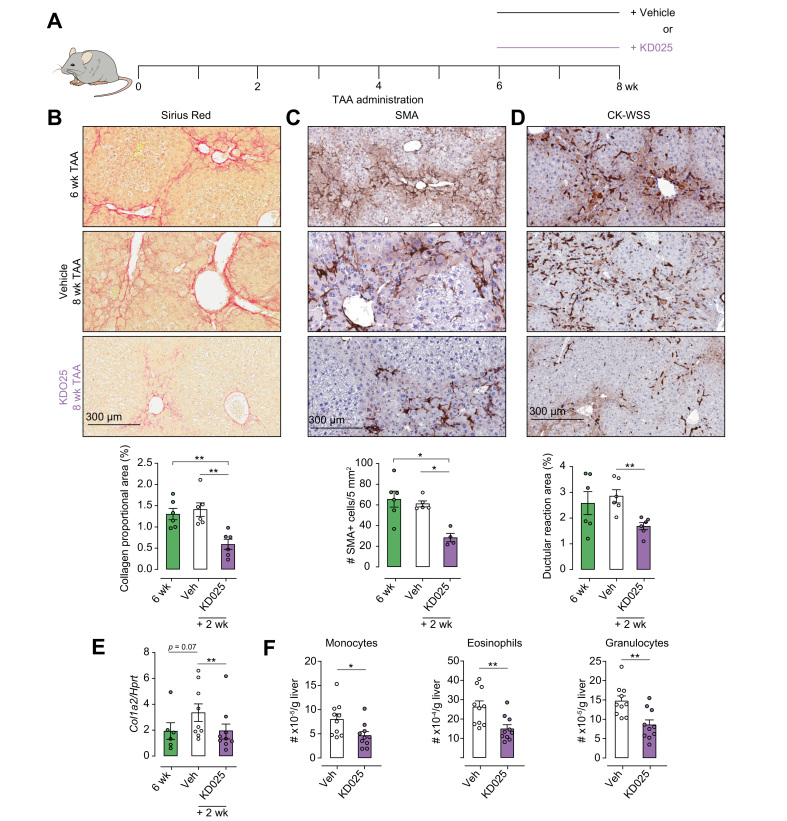
Methods: We used the thioacetamide-induced liver fibrosis model to examine the efficacy of administration of the selective ROCK2 inhibitor KD025 to prevent or treat liver fibrosis and its impact on immune composition and function.
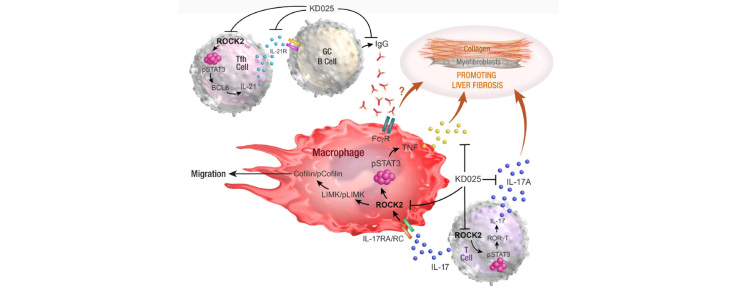
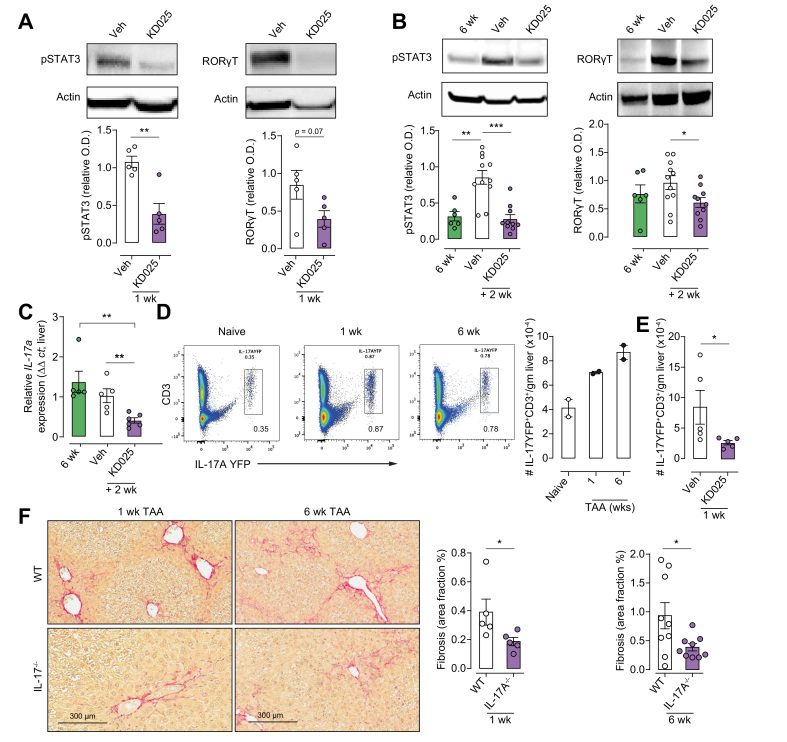
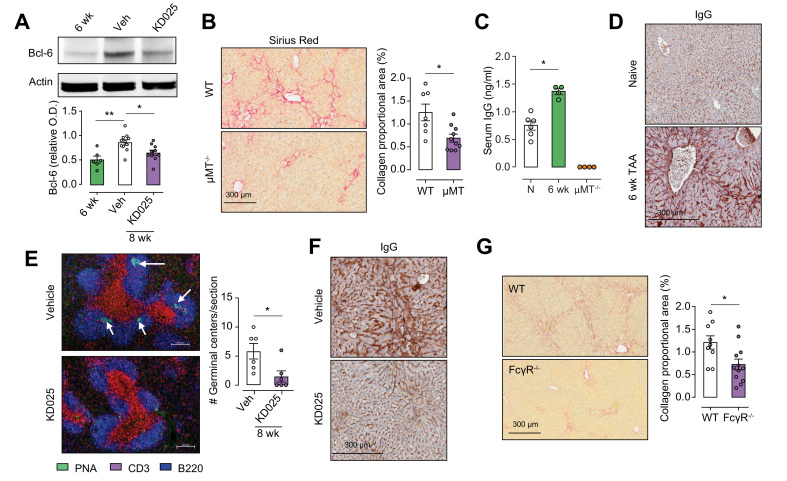
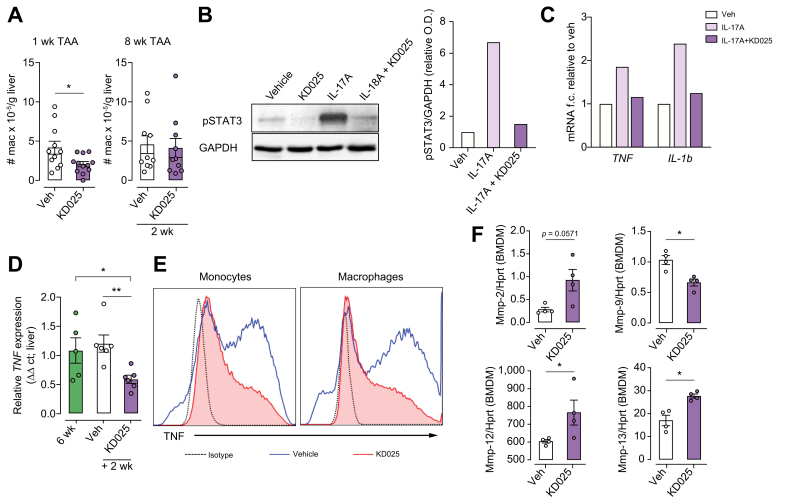
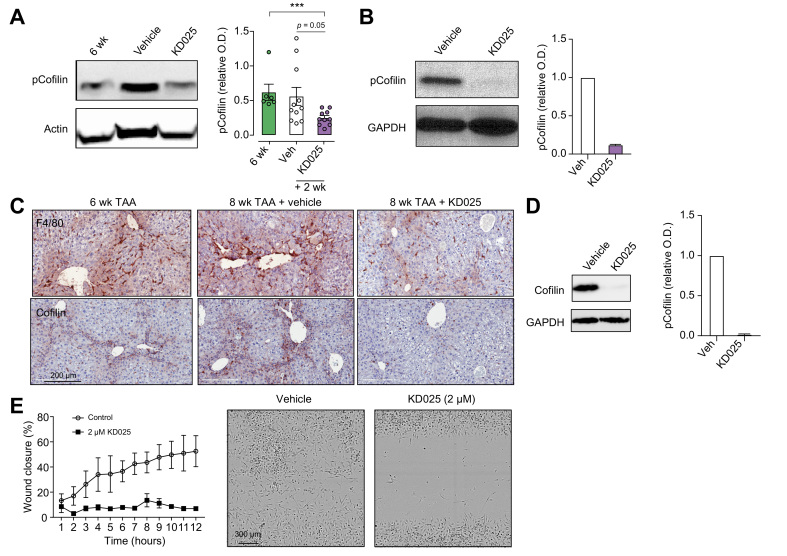
Results: Prophylactic and therapeutic administration of KD025 effectively attenuated thioacetamide-induced liver fibrosis and promoted fibrotic regression. KD025 treatment inhibited liver macrophage tumour necrosis factor production and disrupted the macrophage niche within fibrotic septae. ROCK2 targeting in vitro directly regulated macrophage function through disruption of signal transducer and activator of transcription 3 (STAT3)/cofilin signalling pathways leading to the inhibition of pro-inflammatory cytokine production and macrophage migration. In vivo, KDO25 administration significantly reduced STAT3 phosphorylation and cofilin levels in the liver. Additionally, livers exhibited robust downregulation of immune cell infiltrates and diminished levels of retinoic acid receptor-related orphan receptor gamma (RORγt) and B-cell lymphoma 6 (Bcl6) transcription factors that correlated with a significant reduction in liver IL-17, splenic germinal centre numbers and serum IgG.
Conclusions: As IL-17 and IgG-Fc binding promote pathogenic macrophage differentiation, together our data demonstrate that ROCK2 inhibition prevents and reverses liver fibrosis through direct and indirect effects on macrophage function and highlight the therapeutic potential of ROCK2 inhibition in liver fibrosis.
Lay summary: By using a clinic-ready small-molecule inhibitor, we demonstrate that selective ROCK2 inhibition prevents and reverses hepatic fibrosis through its pleiotropic effects on pro-inflammatory immune cell function. We show that ROCK2 mediates increased IL-17 production, antibody production, and macrophage dysregulation, which together drive fibrogenesis in a model of chemical-induced liver fibrosis. Therefore, in this study, we not only highlight the therapeutic potential of ROCK2 targeting in chronic liver disease but also provide previously undocumented insights into our understanding of cellular and molecular pathways driving the liver fibrosis pathology.
Keywords: ALT, alanine aminotransferase; AST, aspartate aminotransferase; B cells; BMDM, bone marrow-derived macrophages; Bcl6, B-cell lymphoma 6; CLD, chronic liver disease; Col1a2, collagen type α1; DR, ductular reaction; ECM, extracellular matrix; GC, germinal centre; HCC, hepatocellular carcinoma; HSC, hepatic stellate cell; IHC, immunohistochemical; IL-17; Inflammation; LPS, lipopolysaccharide; Liver fibrosis; MMP, matrix metalloproteinase; Macrophages; NASH, non-alcoholic steatohepatitis; RAR, retinoic acid receptor; ROCK, Rho-associated coiled-coil forming protein kinases; ROCK2; ROCK2, Rho-associated kinase 2; RORγt, RAR-related orphan receptor gamma; SR, Sirius red; STAT3, signal transducer and activator of transcription 3; TAA, thioacetamide; TGF-β, transforming growth factor-beta; TNF, tumour necrosis factor; Tfh, T follicular helper; Th17, T helper 17; Therapy; cGVHD, chronic graft-vs-host disease; pCofilin, phosphorylated cofilin; pMac, peritoneal macrophages; pSTAT3, phosphorylated signal transducer and activator of transcription; qRT-PCR, quantitative real-time PCR; α-SMA, alpha smooth muscle actin.
© 2021 The Authors.
Conflict of interest statement
The authors declare no competing financial interests. BRB receives remuneration as an advisor to Magenta Therapeutics and BlueRock Therapeutics; received research funding from BlueRock Therapeutics, Rheos Medicines, Childrens' Cancer Research Fund, and KidsFirst Fund; and is a co-founder of Tmunity. MN, WC, and AZZ are full-time employees of Kadmon Pharmaceuticals, LLC. Please refer to the accompanying ICMJE disclosure forms for further details.
Figures
References
-
- Riento K., Ridley A.J. Rocks: multifunctional kinases in cell behaviour. Nat Rev Mol Cell Biol. 2003;4:446–456. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
