

Proteaphagy is specifically regulated and requires factors dispensable for general autophagy
- PMID: 34919962
- PMCID: PMC8732087
- DOI: 10.1016/j.jbc.2021.101494
Proteaphagy is specifically regulated and requires factors dispensable for general autophagy
Abstract
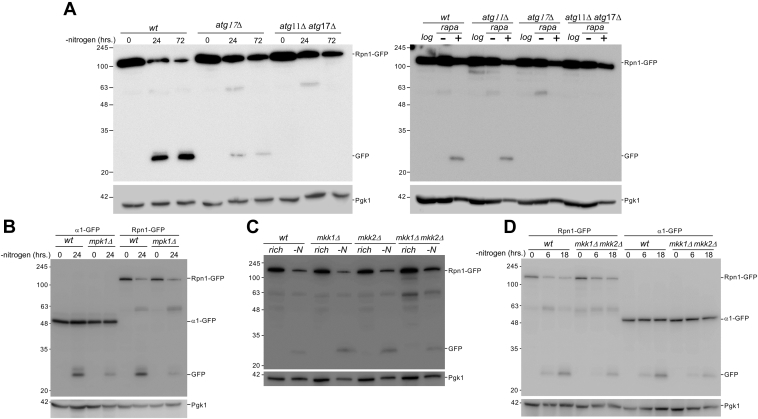
Changing physiological conditions can increase the need for protein degradative capacity in eukaryotic cells. Both the ubiquitin-proteasome system and autophagy contribute to protein degradation. However, these processes can be differently regulated depending on the physiological conditions. Strikingly, proteasomes themselves can be a substrate for autophagy. The signals and molecular mechanisms that govern proteasome autophagy (proteaphagy) are only partly understood. Here, we used immunoblots, native gel analyses, and fluorescent microscopy to understand the regulation of proteaphagy in response to genetic and small molecule-induced perturbations. Our data indicate that chemical inhibition of the master nutrient sensor TORC1 (inhibition of which induces general autophagy) with rapamycin induces a bi-phasic response where proteasome levels are upregulated after an autophagy-dependent reduction. Surprisingly, several conditions that result in inhibited TORC1, such as caffeinine treatment or nitrogen starvation, only induced proteaphagy (i.e., without any proteasome upregulation), suggesting a convergence of signals upstream of proteaphagy under different physiological conditions. Indeed, we found that several conditions that activated general autophagy did not induce proteaphagy, further distinguishing proteaphagy from general autophagy. Consistent with this, we show that Atg11, a selective autophagy receptor, as well as the MAP kinases Mpk1, Mkk1, and Mkk2 all play a role in autophagy of proteasomes, although they are dispensable for general autophagy. Taken together, our data provide new insights into the molecular regulation of proteaphagy by demonstrating that degradation of proteasome complexes is specifically regulated under different autophagy-inducing conditions.
Keywords: autophagy; proteaphagy; proteasome; proteasome inhibitor; protein degradation; starvation; target of rapamycin (TOR); vacuole; yeast.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures



Similar articles
-
The GST-BHMT assay reveals a distinct mechanism underlying proteasome inhibition-induced macroautophagy in mammalian cells.Autophagy. 2015;11(5):812-32. doi: 10.1080/15548627.2015.1034402. Autophagy. 2015. PMID: 25984893 Free PMC article.
-
Mechanisms Regulating the UPS-ALS Crosstalk: The Role of Proteaphagy.Molecules. 2020 May 18;25(10):2352. doi: 10.3390/molecules25102352. Molecules. 2020. PMID: 32443527 Free PMC article. Review.
-
Autophagic Turnover of Inactive 26S Proteasomes in Yeast Is Directed by the Ubiquitin Receptor Cue5 and the Hsp42 Chaperone.Cell Rep. 2016 Aug 9;16(6):1717-1732. doi: 10.1016/j.celrep.2016.07.015. Epub 2016 Jul 28. Cell Rep. 2016. PMID: 27477278
-
Inactive Proteasomes Routed to Autophagic Turnover Are Confined within the Soluble Fraction of the Cell.Biomolecules. 2022 Dec 30;13(1):77. doi: 10.3390/biom13010077. Biomolecules. 2022. PMID: 36671462 Free PMC article.
-
Eat or be eaten: The autophagic plight of inactive 26S proteasomes.Autophagy. 2015;11(10):1927-8. doi: 10.1080/15548627.2015.1078961. Autophagy. 2015. PMID: 26291247 Free PMC article. Review.
Cited by
-
Proteasome assembly chaperone translation upon stress requires Ede1 phase separation at the plasma membrane.iScience. 2023 Dec 14;27(1):108732. doi: 10.1016/j.isci.2023.108732. eCollection 2024 Jan 19. iScience. 2023. PMID: 38235332 Free PMC article.
-
The ribosome-associated chaperone Zuo1 controls translation upon TORC1 inhibition.EMBO J. 2023 Dec 11;42(24):e113240. doi: 10.15252/embj.2022113240. Epub 2023 Nov 20. EMBO J. 2023. PMID: 37984430 Free PMC article.
-
Autophagy-Mediated Cellular Remodeling during Terminal Differentiation of Keratinocytes in the Epidermis and Skin Appendages.Cells. 2024 Oct 10;13(20):1675. doi: 10.3390/cells13201675. Cells. 2024. PMID: 39451193 Free PMC article. Review.
-
Proteasome granule formation is regulated through mitochondrial respiration and kinase signaling.J Cell Sci. 2022 Sep 1;135(17):jcs259778. doi: 10.1242/jcs.259778. Epub 2022 Sep 7. J Cell Sci. 2022. PMID: 35975718 Free PMC article.
-
The Yeast Protein Kinase Sch9 Functions as a Central Nutrient-Responsive Hub That Calibrates Metabolic and Stress-Related Responses.J Fungi (Basel). 2023 Jul 26;9(8):787. doi: 10.3390/jof9080787. J Fungi (Basel). 2023. PMID: 37623558 Free PMC article. Review.
References
-
- Hershko A., Ciechanover A. The ubiquitin system. Annu. Rev. Biochem. 1998;67:425–479. - PubMed
-
- Kraft C., Reggiori F., Peter M. Selective types of autophagy in yeast. Biochim. Biophys. Acta Mol. Cell Res. 2009;1793:1404–1412. - PubMed
-
- Lamb C.A., Dooley H.C., Tooze S.A. Endocytosis and autophagy: Shared machinery for degradation. Bioessays. 2013;35:34–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
