

EpoR stimulates rapid cycling and larger red cells during mouse and human erythropoiesis
- PMID: 34921133
- PMCID: PMC8683474
- DOI: 10.1038/s41467-021-27562-4
EpoR stimulates rapid cycling and larger red cells during mouse and human erythropoiesis
Abstract
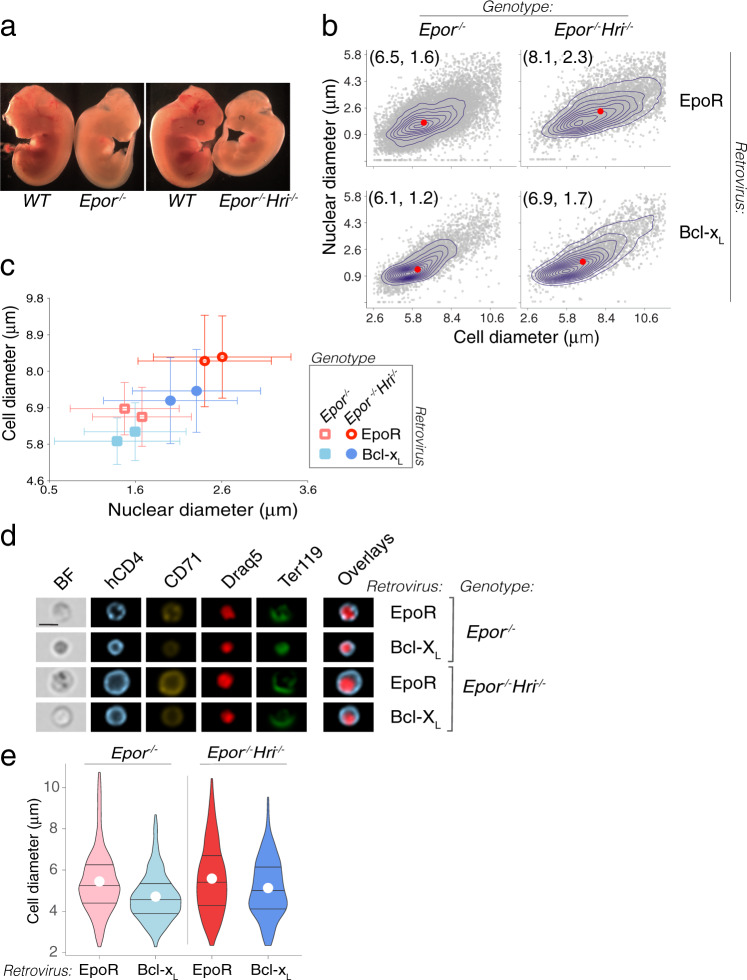
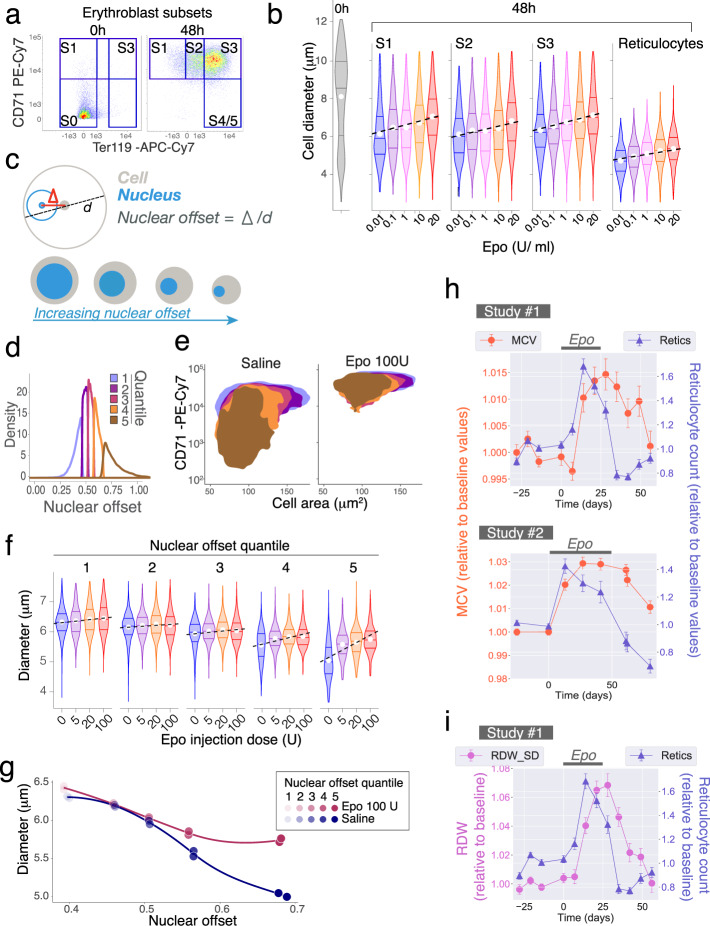
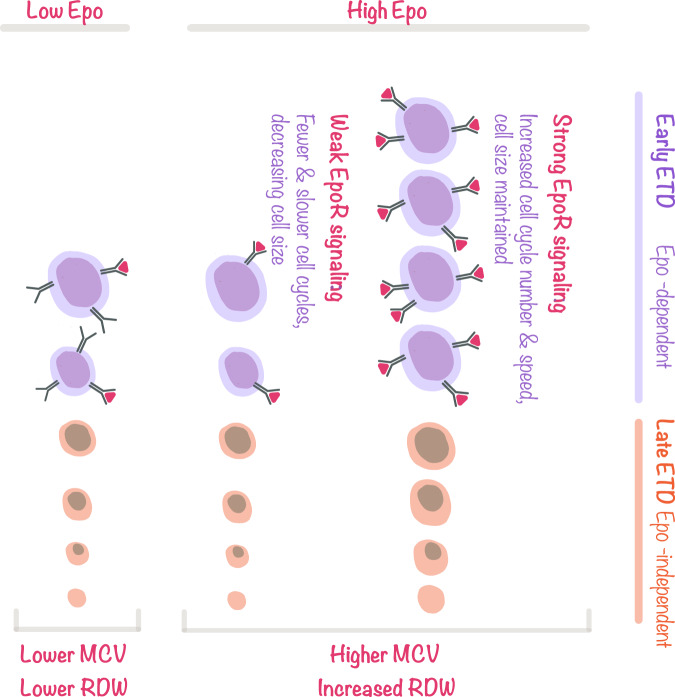
The erythroid terminal differentiation program couples sequential cell divisions with progressive reductions in cell size. The erythropoietin receptor (EpoR) is essential for erythroblast survival, but its other functions are not well characterized. Here we use Epor-/- mouse erythroblasts endowed with survival signaling to identify novel non-redundant EpoR functions. We find that, paradoxically, EpoR signaling increases red cell size while also increasing the number and speed of erythroblast cell cycles. EpoR-regulation of cell size is independent of established red cell size regulation by iron. High erythropoietin (Epo) increases red cell size in wild-type mice and in human volunteers. The increase in mean corpuscular volume (MCV) outlasts the duration of Epo treatment and is not the result of increased reticulocyte number. Our work shows that EpoR signaling alters the relationship between cycling and cell size. Further, diagnostic interpretations of increased MCV should now include high Epo levels and hypoxic stress.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
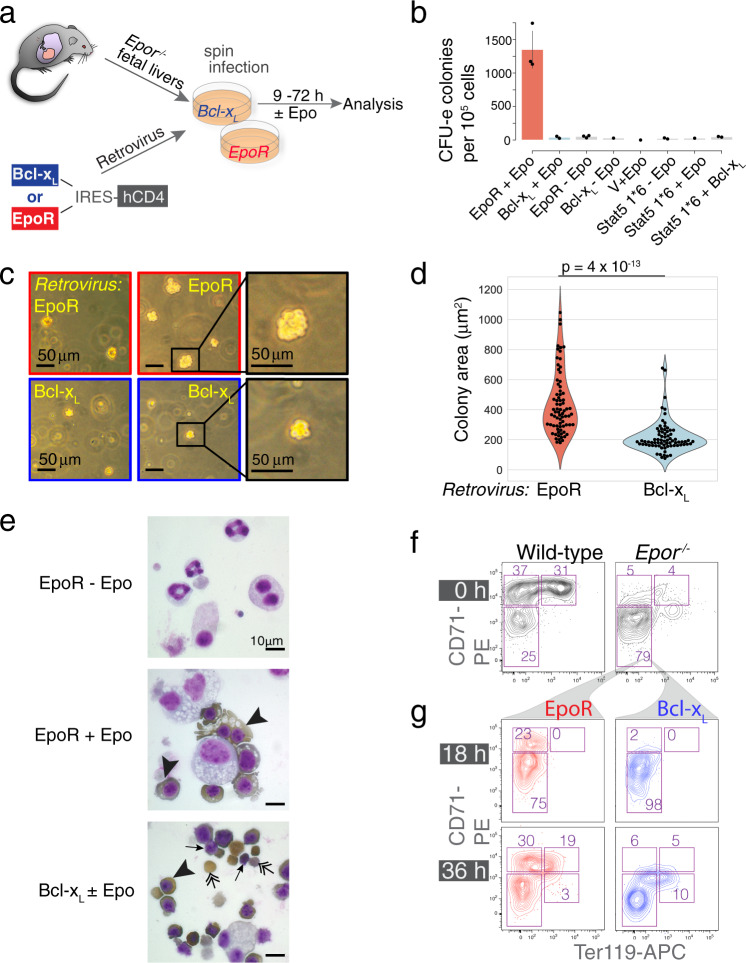
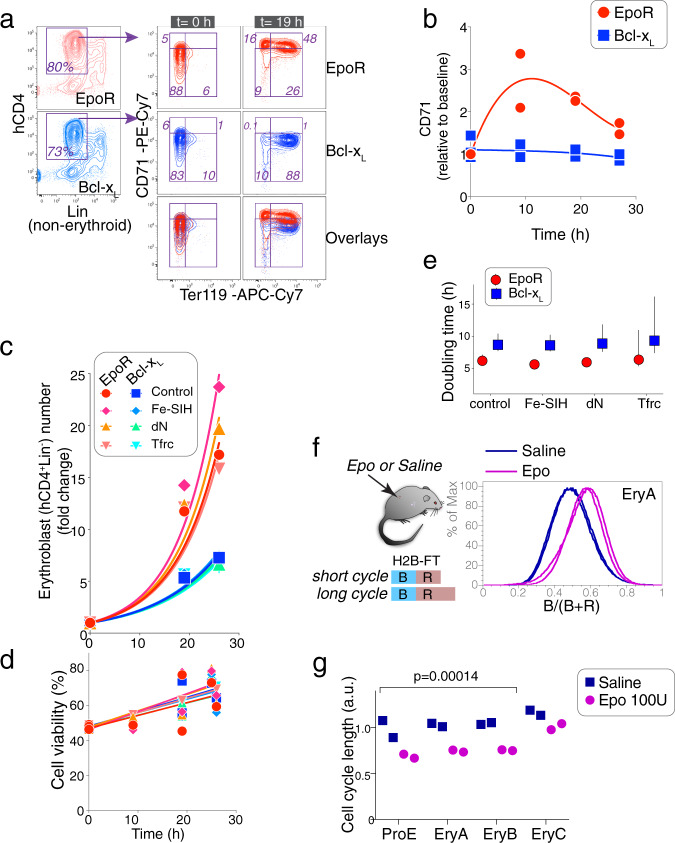
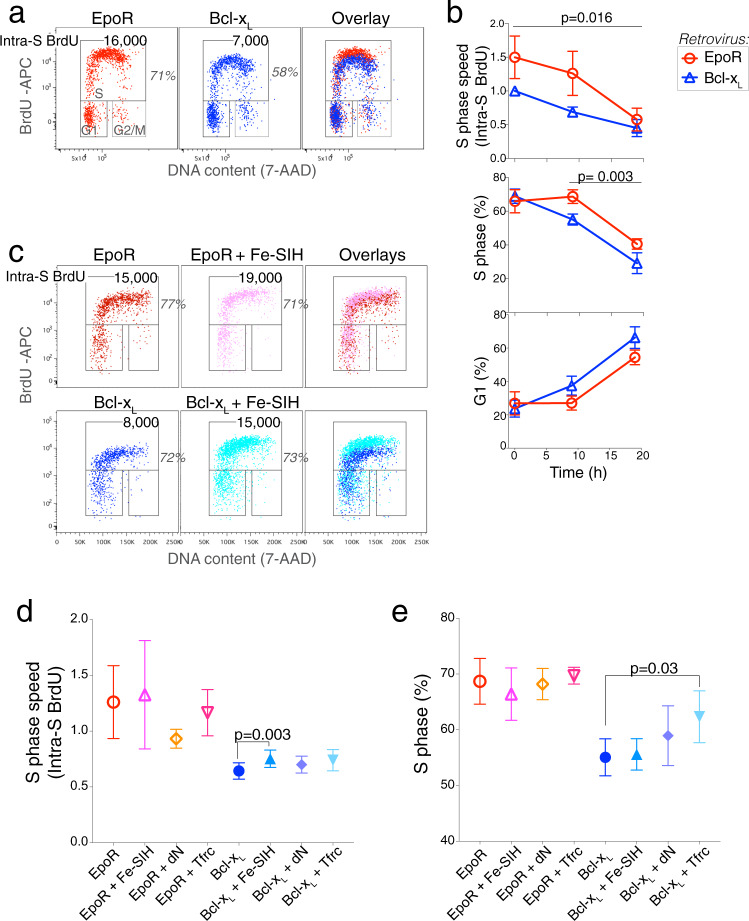
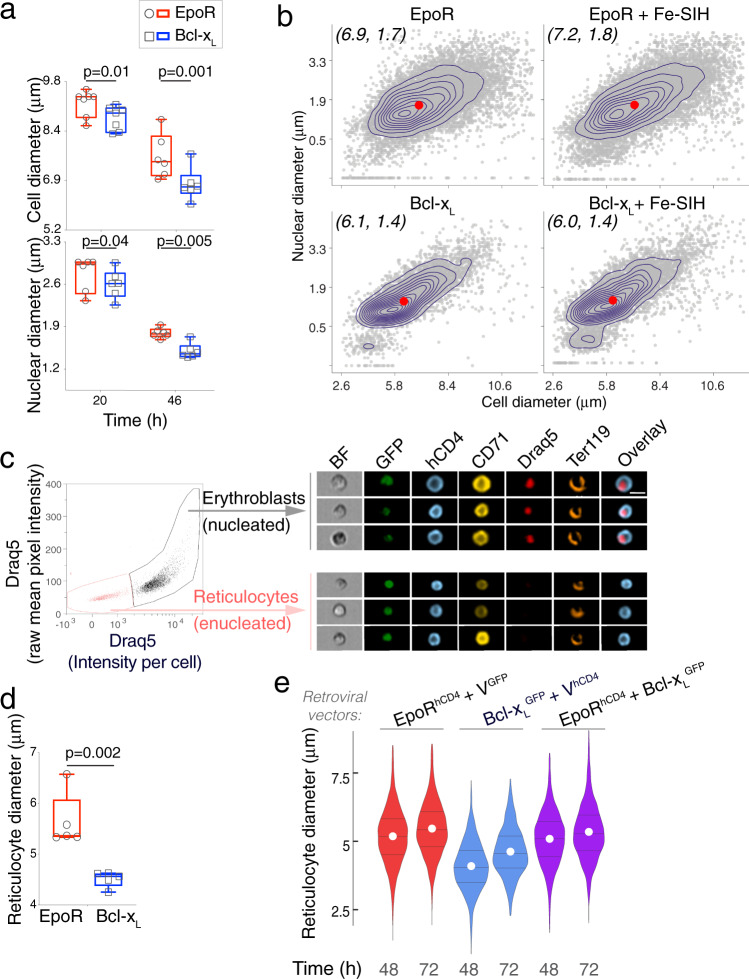
References
-
- D’Andrea AD, Fasman GD, Lodish HF. Erythropoietin receptor and interleukin-2 receptor b chain: a new receptor family. Cell. 1989;58:1023–1024. - PubMed
-
- Wu H, Liu X, Jaenisch R, Lodish HF. Generation of committed erythroid BFU-E and CFU-E progenitors does not require erythropoietin or the erythropoietin receptor. Cell. 1995;83:59–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
