

The plasma membrane H+ -ATPase FgPMA1 regulates the development, pathogenicity, and phenamacril sensitivity of Fusarium graminearum by interacting with FgMyo-5 and FgBmh2
- PMID: 34921490
- PMCID: PMC8916210
- DOI: 10.1111/mpp.13173
The plasma membrane H+ -ATPase FgPMA1 regulates the development, pathogenicity, and phenamacril sensitivity of Fusarium graminearum by interacting with FgMyo-5 and FgBmh2
Abstract
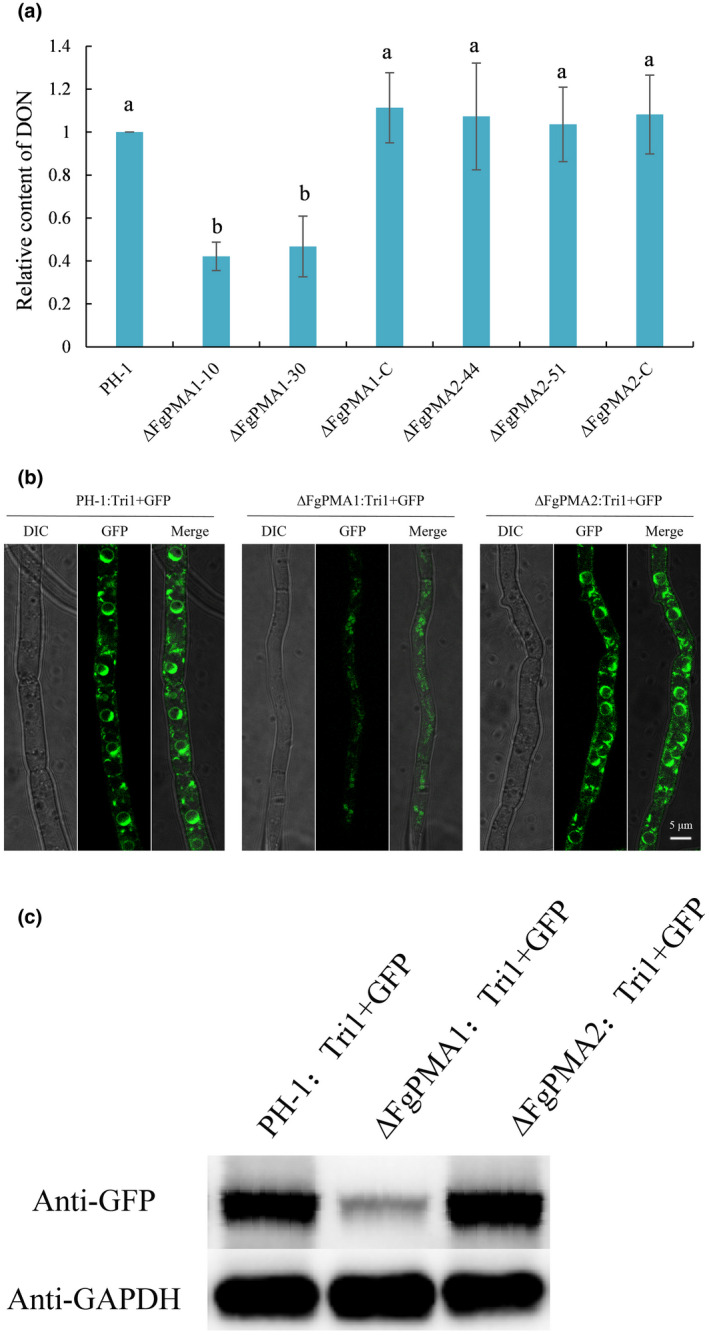
Fusarium graminearum, as the causal agent of Fusarium head blight (FHB), not only causes yield loss, but also contaminates the quality of wheat by producing mycotoxins, such as deoxynivalenol (DON). The plasma membrane H+ -ATPases play important roles in many growth stages in plants and yeasts, but their functions and regulation in phytopathogenic fungi remain largely unknown. Here we characterized two plasma membrane H+ -ATPases: FgPMA1 and FgPMA2 in F. graminearum. The FgPMA1 deletion mutant (∆FgPMA1), but not FgPMA2 deletion mutant (∆FgPMA2), was impaired in vegetative growth, pathogenicity, and sexual and asexual development. FgPMA1 was localized to the plasma membrane, and ∆FgPMA1 displayed reduced integrity of plasma membrane. ∆FgPMA1 not only impaired the formation of the toxisome, which is a compartment where DON is produced, but also suppressed the expression level of DON biosynthetic enzymes, decreased DON production, and decreased the amount of mycelial invasion, leading to impaired pathogenicity by exclusively developing disease on inoculation sites of wheat ears and coleoptiles. ∆FgPMA1 exhibited decreased sensitivity to some osmotic stresses, a cell wall-damaging agent (Congo red), a cell membrane-damaging agent (sodium dodecyl sulphate), and heat shock stress. FgMyo-5 is the target of phenamacril used for controlling FHB. We found FgPMA1 interacted with FgMyo-5, and ∆FgPMA1 showed an increased expression level of FgMyo-5, resulting in increased sensitivity to phenamacril, but not to other fungicides. Furthermore, co-immunoprecipitation confirmed that FgPMA1, FgMyo-5, and FgBmh2 (a 14-3-3 protein) form a complex to regulate the sensitivity to phenamacril and biological functions. Collectively, this study identified a novel regulating mechanism of FgPMA1 in pathogenicity and phenamacril sensitivity of F. graminearum.
Keywords: Fusarium graminearum; DON production; FgPMA1; pathogenicity; phenamacril.
© 2021 The Authors. Molecular Plant Pathology published by British Society for Plant Pathology and John Wiley & Sons Ltd.
Figures







Similar articles
-
The b-ZIP transcription factor FgTfmI is required for the fungicide phenamacril tolerance and pathogenecity in Fusarium graminearum.Pest Manag Sci. 2019 Dec;75(12):3312-3322. doi: 10.1002/ps.5454. Epub 2019 Jun 3. Pest Manag Sci. 2019. PMID: 31025482
-
Characterization of the fludioxonil and phenamacril dual resistant mutants of Fusarium graminearum.Pestic Biochem Physiol. 2024 Mar;200:105815. doi: 10.1016/j.pestbp.2024.105815. Epub 2024 Feb 5. Pestic Biochem Physiol. 2024. PMID: 38582573
-
Activity of Demethylation Inhibitor Fungicide Metconazole on Chinese Fusarium graminearum Species Complex and Its Application in Carbendazim-Resistance Management of Fusarium Head Blight in Wheat.Plant Dis. 2019 May;103(5):929-937. doi: 10.1094/PDIS-09-18-1592-RE. Epub 2019 Mar 18. Plant Dis. 2019. PMID: 30880557
-
Contribution of cell wall degrading enzymes to pathogenesis of Fusarium graminearum: a review.J Basic Microbiol. 2009 Jun;49(3):231-41. doi: 10.1002/jobm.200800231. J Basic Microbiol. 2009. PMID: 19025875 Review.
-
Comprehensive analysis of the mechanisms conferring resistance to phenamacril in the Fusarium species.Front Cell Infect Microbiol. 2025 Feb 11;15:1536532. doi: 10.3389/fcimb.2025.1536532. eCollection 2025. Front Cell Infect Microbiol. 2025. PMID: 40007612 Free PMC article. Review.
Cited by
-
FgMet3 and FgMet14 related to cysteine and methionine biosynthesis regulate vegetative growth, sexual reproduction, pathogenicity, and sensitivity to fungicides in Fusarium graminearum.Front Plant Sci. 2022 Oct 24;13:1011709. doi: 10.3389/fpls.2022.1011709. eCollection 2022. Front Plant Sci. 2022. PMID: 36352883 Free PMC article.
-
Redundant and Distinct Roles of Two 14-3-3 Proteins in Fusarium sacchari, Pathogen of Sugarcane Pokkah Boeng Disease.J Fungi (Basel). 2024 Mar 28;10(4):257. doi: 10.3390/jof10040257. J Fungi (Basel). 2024. PMID: 38667928 Free PMC article.
-
FgPfn participates in vegetative growth, sexual reproduction, pathogenicity, and fungicides sensitivity via affecting both microtubules and actin in the filamentous fungus Fusarium graminearum.PLoS Pathog. 2024 May 3;20(5):e1012215. doi: 10.1371/journal.ppat.1012215. eCollection 2024 May. PLoS Pathog. 2024. PMID: 38701108 Free PMC article.
-
The Plasma Membrane H+ ATPase CsPMA2 Regulates Lipid Droplet Formation, Appressorial Development and Virulence in Colletotrichum siamense.Int J Mol Sci. 2023 Dec 11;24(24):17337. doi: 10.3390/ijms242417337. Int J Mol Sci. 2023. PMID: 38139168 Free PMC article.
-
Transcription Activator FgDDT Interacts With FgISW1 to Regulate Fungal Development and Pathogenicity in the Global Pathogen Fusarium graminearum.Mol Plant Pathol. 2025 Apr;26(4):e70076. doi: 10.1111/mpp.70076. Mol Plant Pathol. 2025. PMID: 40151047 Free PMC article.
References
-
- Addison, R. (1986) Primary structure of the Neurospora plasma membrane H+‐ATPase deduced from the gene sequence‐homology to Na+/K+‐, Ca2+‐, and K+‐ATPase. Journal of Biological Chemistry, 261, 4896–4901. - PubMed
-
- Alexander, N.J. , Proctor, R.H. & McCormick, S.P. (2009) Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium . Toxin Reviews, 28, 198–215.
-
- Ambesi, A. , Miranda, M. , Petrov, V.V. & Slayman, C.W. (2000) Biogenesis and function of the yeast plasma‐membrane H+‐ATPase. Joural of Experimental Biology, 203, 155–160. - PubMed
-
- Baunsgaard, L. , Venema, K. , Axelsen, K.B. , Villalba, J.M. , Welling, A. , Wollenweber, B. et al. (1996) Modified plant plasma membrane H+‐ATPase with improved transport coupling efficiency identified by mutant selection in yeast. The Plant Journal, 10, 451–458. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
