

Dynamic changes in the association between maternal mRNAs and endoplasmic reticulum during ascidian early embryogenesis
- PMID: 34921621
- PMCID: PMC8918112
- DOI: 10.1007/s00427-021-00683-y
Dynamic changes in the association between maternal mRNAs and endoplasmic reticulum during ascidian early embryogenesis
Abstract
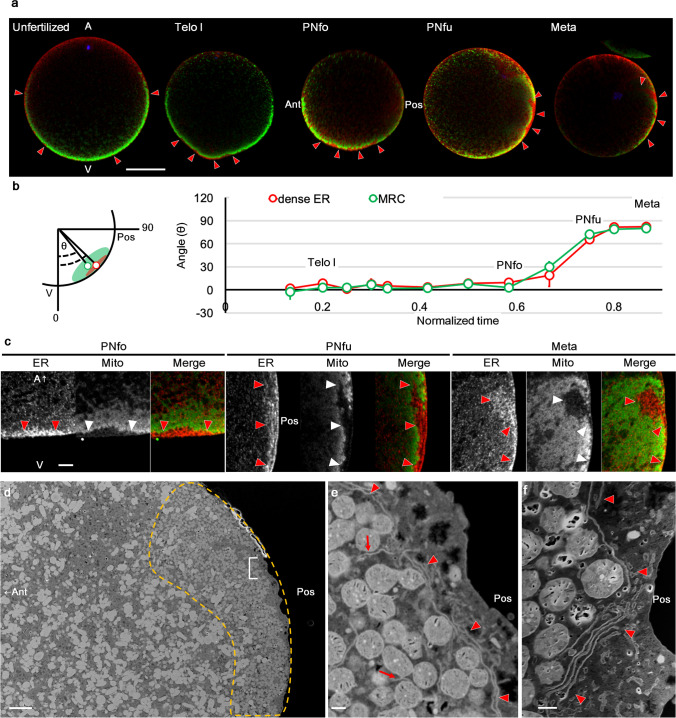
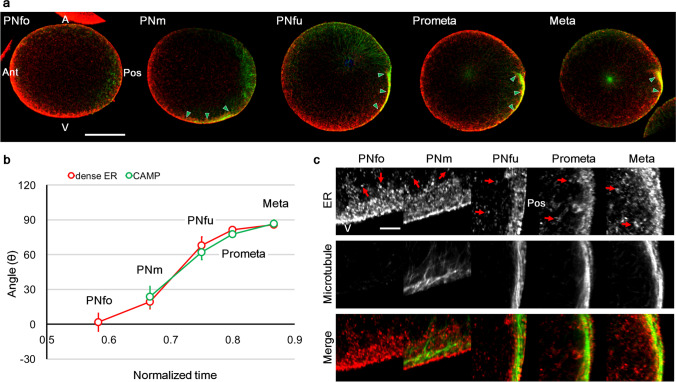
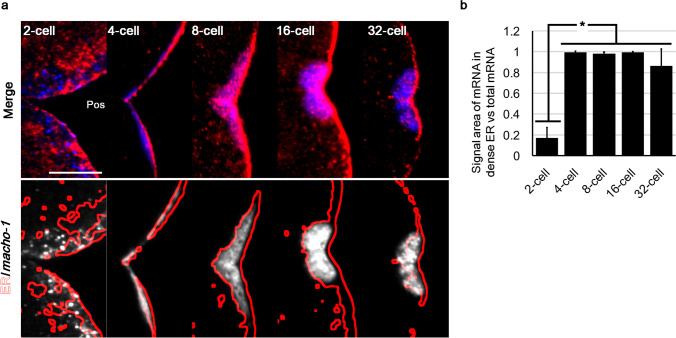
Axis formation is one of the most important events occurring at the beginning of animal development. In the ascidian egg, the antero-posterior axis is established at this time owing to a dynamic cytoplasmic movement called cytoplasmic and cortical reorganisation. During this movement, mitochondria, endoplasmic reticulum (ER), and maternal mRNAs (postplasmic/PEM RNAs) are translocated to the future posterior side. Although accumulating evidence indicates the crucial roles played by the asymmetrical localisation of these organelles and the translational regulation of postplasmic/PEM RNAs, the organisation of ER has not been described in sufficient detail to date owing to technical difficulties. In this study, we developed three different multiple staining protocols for visualising the ER in combination with mitochondria, microtubules, or mRNAs in whole-mount specimens. We defined the internally expanded "dense ER" using these protocols and described cisterna-like structures of the dense ER using focused ion beam-scanning electron microscopy. Most importantly, we described the dynamic changes in the colocalisation of postplasmic/PEM mRNAs and dense ER; for example, macho-1 mRNA was detached and excluded from the dense ER during the second phase of ooplasmic movements. These detailed descriptions of the association between maternal mRNA and ER can provide clues for understanding the translational regulation mechanisms underlying axis determination during ascidian early embryogenesis.
Keywords: Axis determination; Cytoskeleton; ER translocation; Maternal mRNA; Translational regulation.
© 2021. The Author(s).
Figures



Similar articles
-
Cytoplasmic localization and reorganization in ascidian eggs: role of postplasmic/PEM RNAs in axis formation and fate determination.Wiley Interdiscip Rev Dev Biol. 2012 Jul-Aug;1(4):501-18. doi: 10.1002/wdev.54. Epub 2012 Apr 9. Wiley Interdiscip Rev Dev Biol. 2012. PMID: 23801532 Review.
-
Maternal determinants and mRNAs in the cortex of ascidian oocytes, zygotes and embryos.Biol Cell. 2005 Jan;97(1):35-49. doi: 10.1042/BC20040126. Biol Cell. 2005. PMID: 15601256 Review.
-
POPK-1/Sad-1 kinase is required for the proper translocation of maternal mRNAs and putative germ plasm at the posterior pole of the ascidian embryo.Development. 2005 Nov;132(21):4731-42. doi: 10.1242/dev.02049. Epub 2005 Oct 5. Development. 2005. PMID: 16207760
-
Cortical anchorages and cell type segregations of maternal postplasmic/PEM RNAs in ascidians.Dev Biol. 2009 Dec 1;336(1):96-111. doi: 10.1016/j.ydbio.2009.09.001. Epub 2009 Sep 6. Dev Biol. 2009. PMID: 19735652
-
From oocyte to 16-cell stage: cytoplasmic and cortical reorganizations that pattern the ascidian embryo.Dev Dyn. 2007 Jul;236(7):1716-31. doi: 10.1002/dvdy.21136. Dev Dyn. 2007. PMID: 17420986 Review.
Cited by
-
Morphological Evidence for Novel Roles of Microtubules in Macrophage Phagocytosis.Int J Mol Sci. 2023 Jan 10;24(2):1373. doi: 10.3390/ijms24021373. Int J Mol Sci. 2023. PMID: 36674886 Free PMC article.
-
Actin Filament in the First Cell Cycle Contributes to the Determination of the Anteroposterior Axis in Ascidian Development.J Dev Biol. 2022 Feb 4;10(1):10. doi: 10.3390/jdb10010010. J Dev Biol. 2022. PMID: 35225963 Free PMC article.
References
-
- Conklin EG. The organization and cell lineage of the ascidian egg. J Acad Nat Sci Phila. 1905;13:1–119.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
