

Subcellular Transcriptomics and Proteomics: A Comparative Methods Review
- PMID: 34922010
- PMCID: PMC8864473
- DOI: 10.1016/j.mcpro.2021.100186
Subcellular Transcriptomics and Proteomics: A Comparative Methods Review
Abstract
The internal environment of cells is molecularly crowded, which requires spatial organization via subcellular compartmentalization. These compartments harbor specific conditions for molecules to perform their biological functions, such as coordination of the cell cycle, cell survival, and growth. This compartmentalization is also not static, with molecules trafficking between these subcellular neighborhoods to carry out their functions. For example, some biomolecules are multifunctional, requiring an environment with differing conditions or interacting partners, and others traffic to export such molecules. Aberrant localization of proteins or RNA species has been linked to many pathological conditions, such as neurological, cancer, and pulmonary diseases. Differential expression studies in transcriptomics and proteomics are relatively common, but the majority have overlooked the importance of subcellular information. In addition, subcellular transcriptomics and proteomics data do not always colocate because of the biochemical processes that occur during and after translation, highlighting the complementary nature of these fields. In this review, we discuss and directly compare the current methods in spatial proteomics and transcriptomics, which include sequencing- and imaging-based strategies, to give the reader an overview of the current tools available. We also discuss current limitations of these strategies as well as future developments in the field of spatial -omics.
Keywords: cellular fractionation; imaging; proximity labeling; spatial proteomics; spatial transcriptomics.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare no competing interests.
Figures
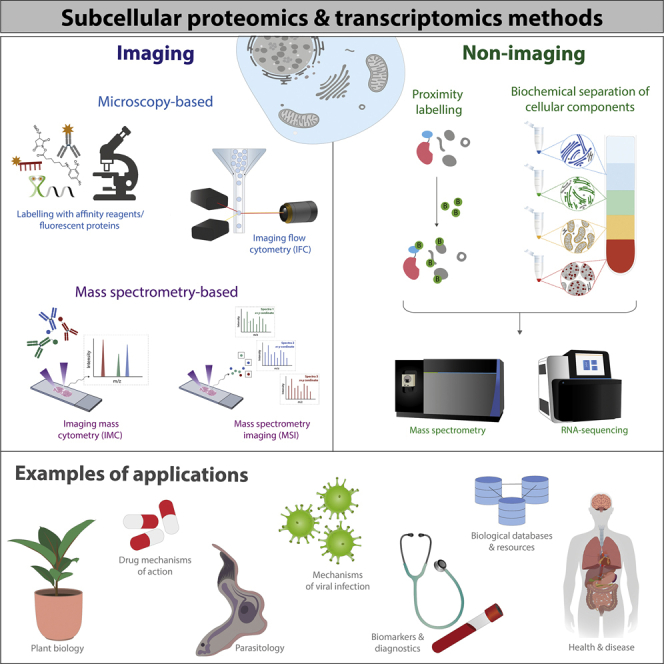
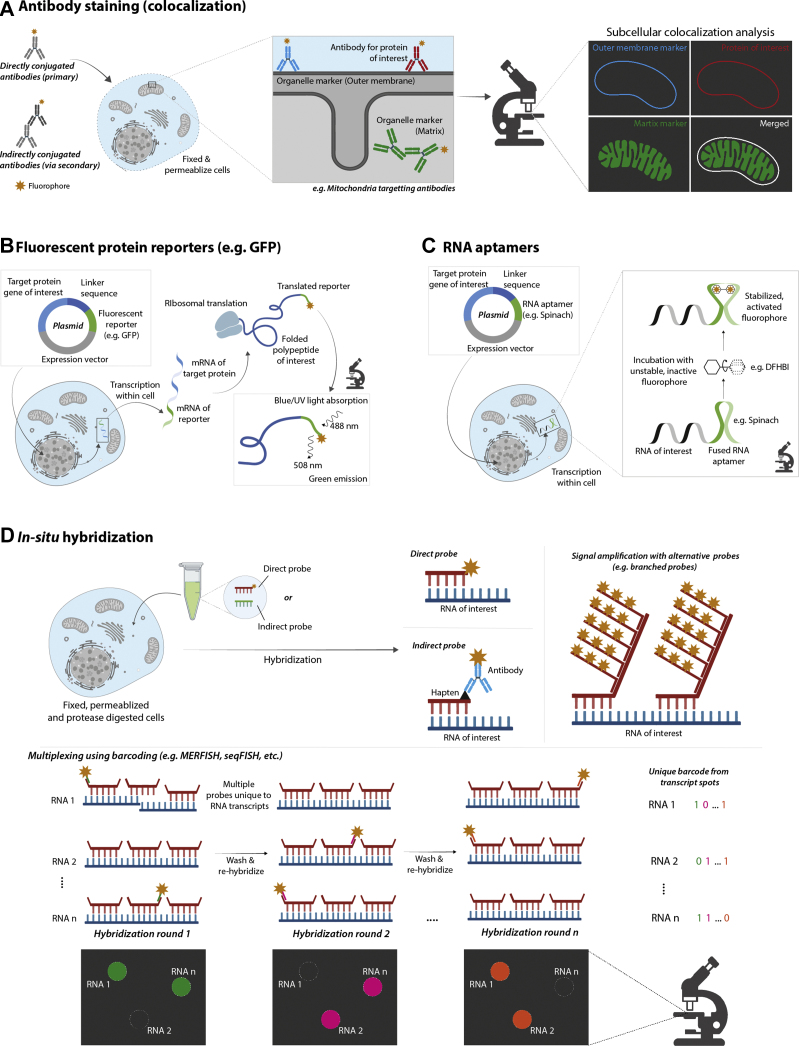
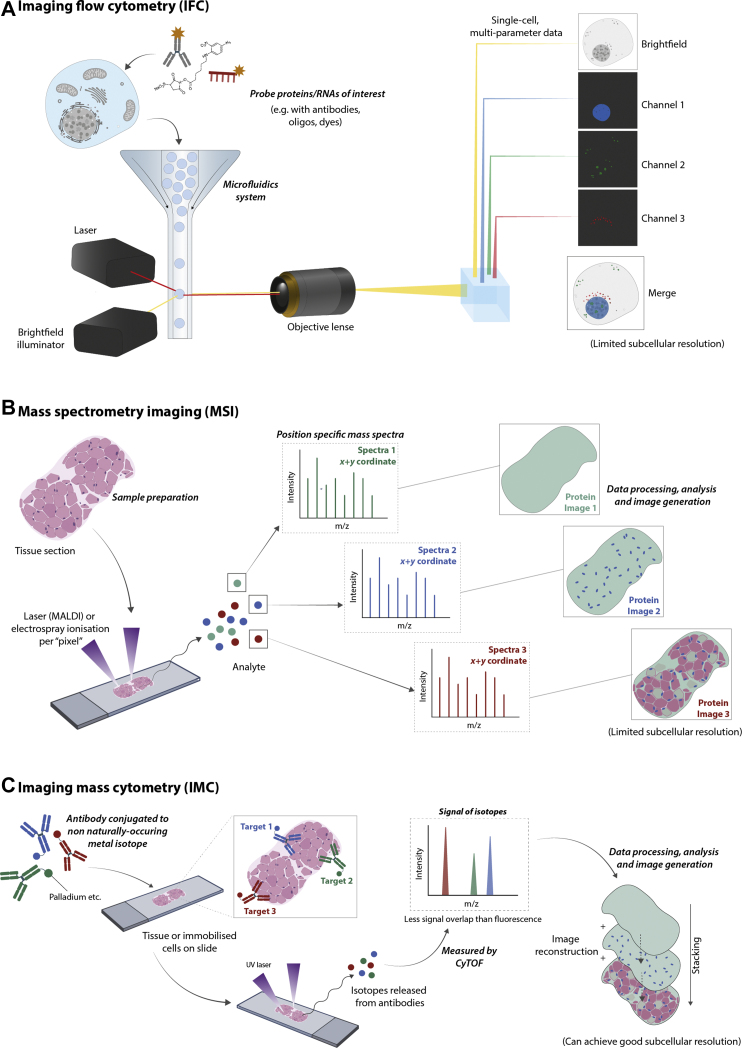
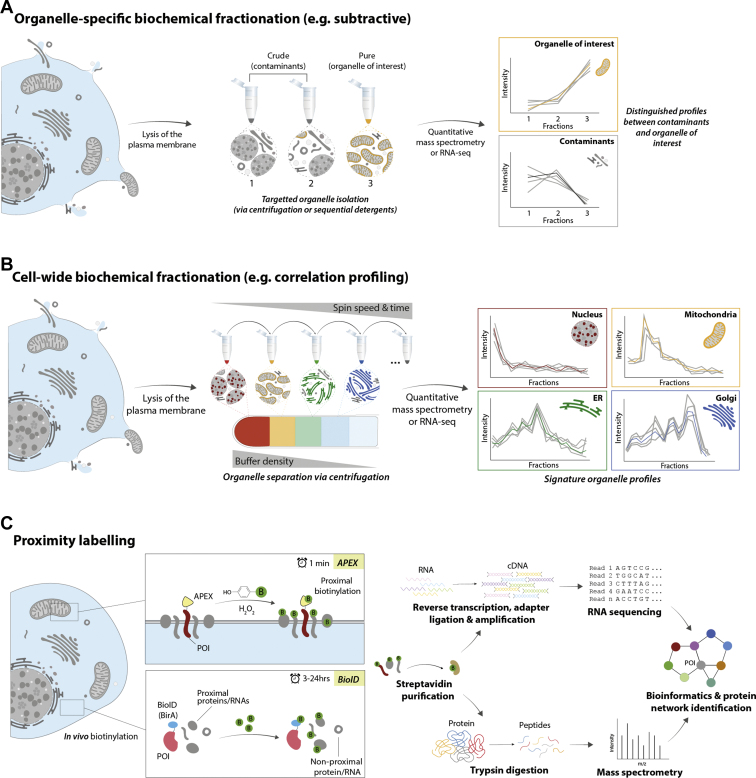
Similar articles
-
Microbial Pathogenesis in the Era of Spatial Omics.Infect Immun. 2023 Jul 18;91(7):e0044222. doi: 10.1128/iai.00442-22. Epub 2023 May 31. Infect Immun. 2023. PMID: 37255461 Free PMC article. Review.
-
Spatial perspectives in the redox code-Mass spectrometric proteomics studies of moonlighting proteins.Mass Spectrom Rev. 2018 Jan;37(1):81-100. doi: 10.1002/mas.21508. Epub 2016 May 17. Mass Spectrom Rev. 2018. PMID: 27186965 Review.
-
Computational solutions for spatial transcriptomics.Comput Struct Biotechnol J. 2022 Sep 1;20:4870-4884. doi: 10.1016/j.csbj.2022.08.043. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36147664 Free PMC article. Review.
-
Single cell- and spatial 'Omics revolutionize physiology.Acta Physiol (Oxf). 2022 Aug;235(4):e13848. doi: 10.1111/apha.13848. Epub 2022 Jun 14. Acta Physiol (Oxf). 2022. PMID: 35656634 Review.
-
Subcellular fractionation methods and strategies for proteomics.Proteomics. 2010 Nov;10(22):3935-56. doi: 10.1002/pmic.201000289. Epub 2010 Nov 2. Proteomics. 2010. PMID: 21080488 Review.
Cited by
-
Proximitomics by Reactive Species.ACS Cent Sci. 2024 Jun 12;10(6):1135-1147. doi: 10.1021/acscentsci.4c00373. eCollection 2024 Jun 26. ACS Cent Sci. 2024. PMID: 38947200 Free PMC article. Review.
-
Unveiling spatial complexity in solid tumor immune microenvironments through multiplexed imaging.Front Immunol. 2024 Mar 19;15:1383932. doi: 10.3389/fimmu.2024.1383932. eCollection 2024. Front Immunol. 2024. PMID: 38566984 Free PMC article.
-
Proximity Labeling in Plants.Annu Rev Plant Biol. 2023 May 22;74:285-312. doi: 10.1146/annurev-arplant-070522-052132. Epub 2023 Feb 28. Annu Rev Plant Biol. 2023. PMID: 36854476 Free PMC article. Review.
-
The family of glutathione peroxidase proteins and their role against biotic stress in plants: a systematic review.Front Plant Sci. 2025 Feb 20;16:1425880. doi: 10.3389/fpls.2025.1425880. eCollection 2025. Front Plant Sci. 2025. PMID: 40051871 Free PMC article.
-
Dual-Signal Feature Spaces Map Protein Subcellular Locations Based on Immunohistochemistry Image and Protein Sequence.Sensors (Basel). 2023 Nov 7;23(22):9014. doi: 10.3390/s23229014. Sensors (Basel). 2023. PMID: 38005402 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
