

Common sequence motifs of nascent chains engage the ribosome surface and trigger factor
- PMID: 34930833
- PMCID: PMC8719866
- DOI: 10.1073/pnas.2103015118
Common sequence motifs of nascent chains engage the ribosome surface and trigger factor
Abstract
In the cell, the conformations of nascent polypeptide chains during translation are modulated by both the ribosome and its associated molecular chaperone, trigger factor. The specific interactions that underlie these modulations, however, are still not known in detail. Here, we combine protein engineering, in-cell and in vitro NMR spectroscopy, and molecular dynamics simulations to explore how proteins interact with the ribosome during their biosynthesis before folding occurs. Our observations of α-synuclein nascent chains in living Escherichia coli cells reveal that ribosome surface interactions dictate the dynamics of emerging disordered polypeptides in the crowded cytosol. We show that specific basic and aromatic motifs drive such interactions and directly compete with trigger factor binding while biasing the direction of the nascent chain during its exit out of the tunnel. These results reveal a structural basis for the functional role of the ribosome as a scaffold with holdase characteristics and explain how handover of the nascent chain to specific auxiliary proteins occurs among a host of other factors in the cytosol.
Keywords: NMR spectroscopy; alpha synuclein; cotranslational folding; in-cell NMR; structural biology.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
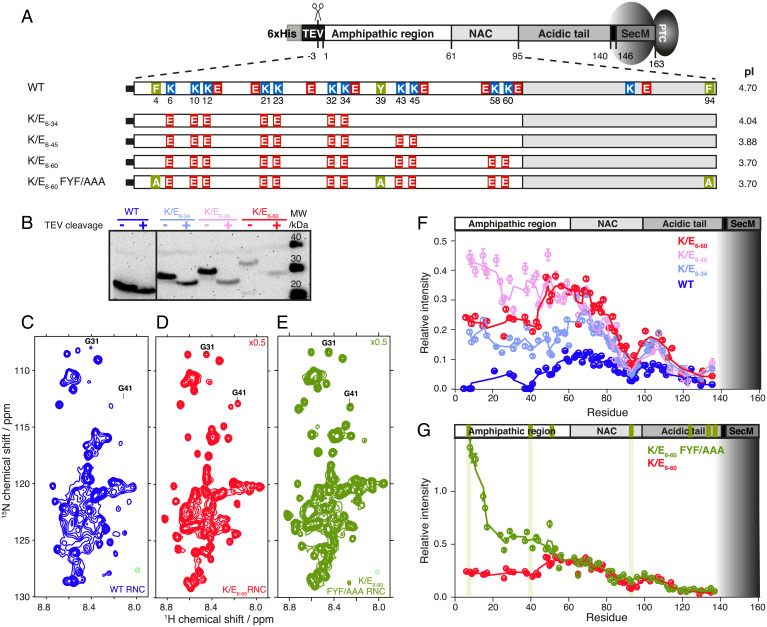
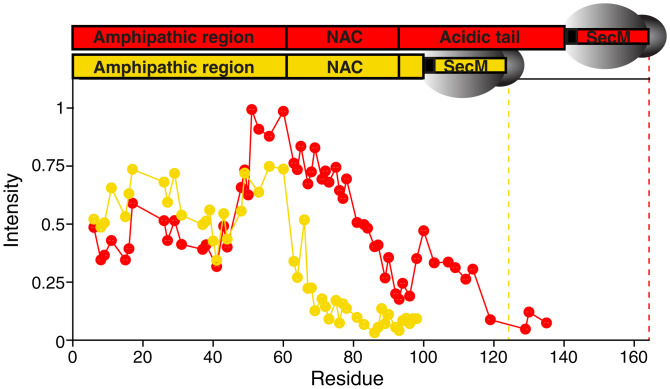
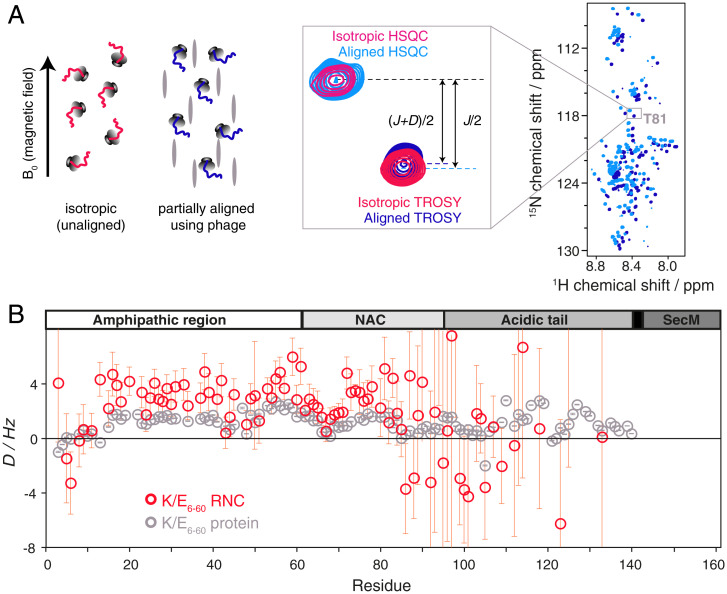
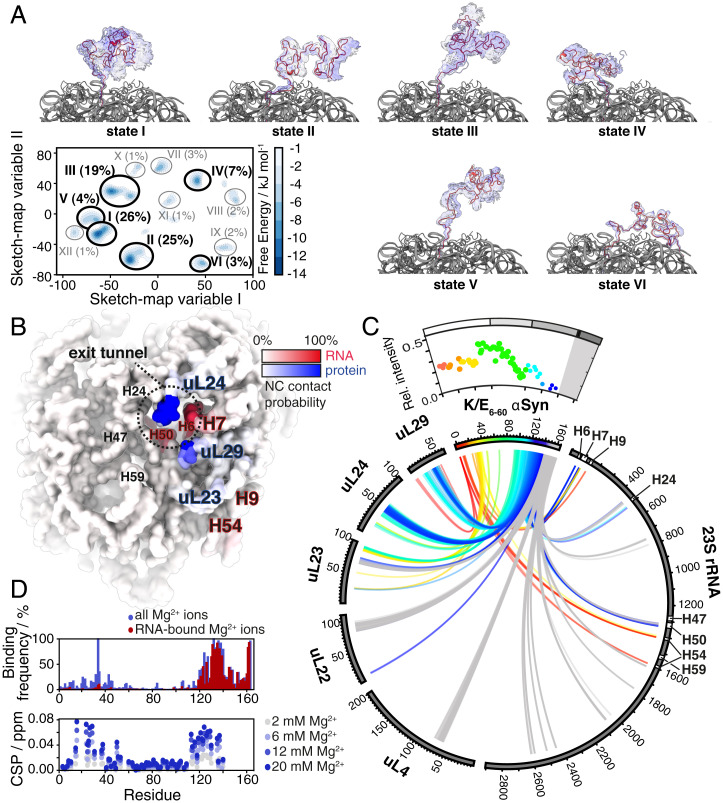
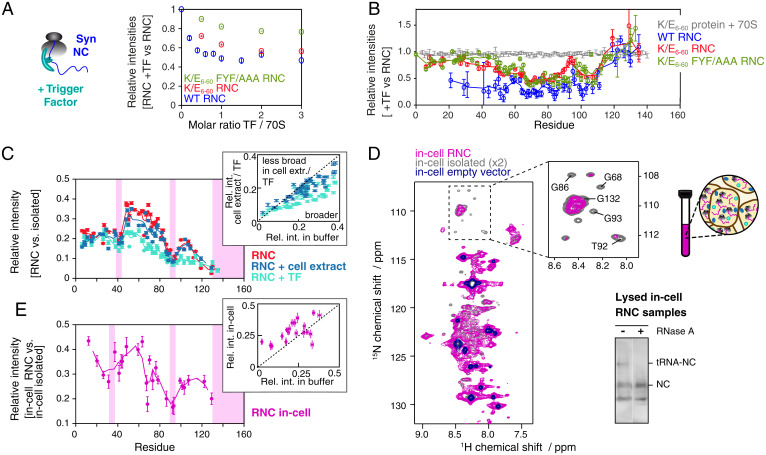
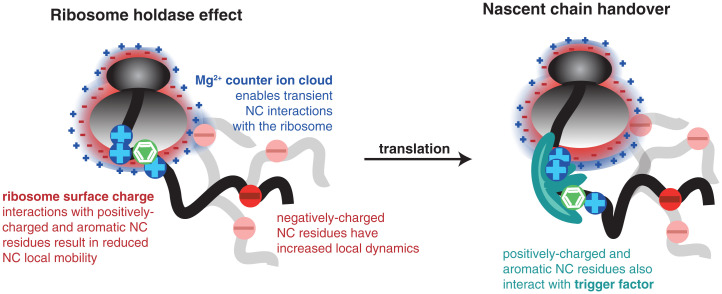
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
