

Microbial metabolite delta-valerobetaine is a diet-dependent obesogen
- PMID: 34931082
- PMCID: PMC8711632
- DOI: 10.1038/s42255-021-00502-8
Microbial metabolite delta-valerobetaine is a diet-dependent obesogen
Abstract
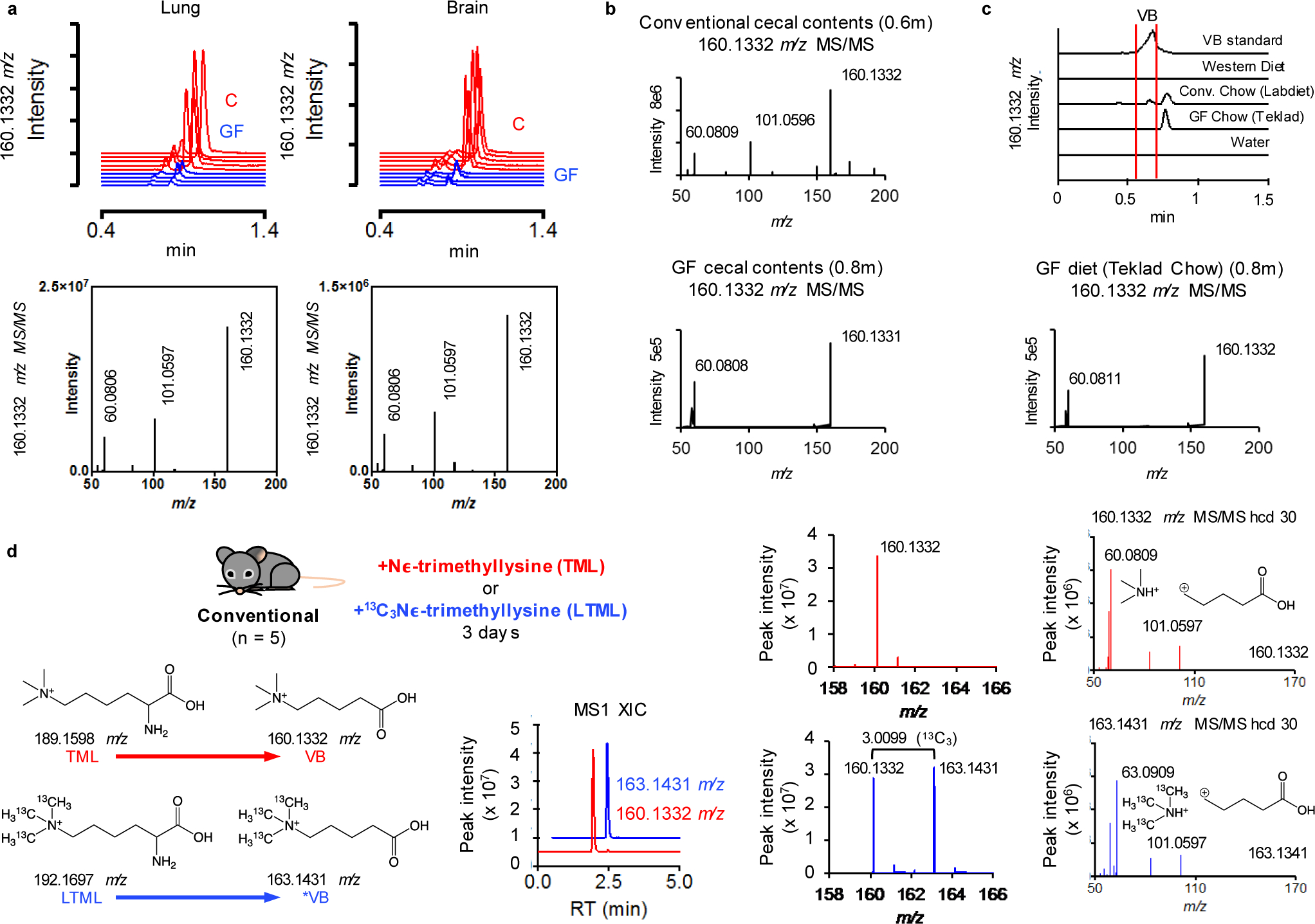
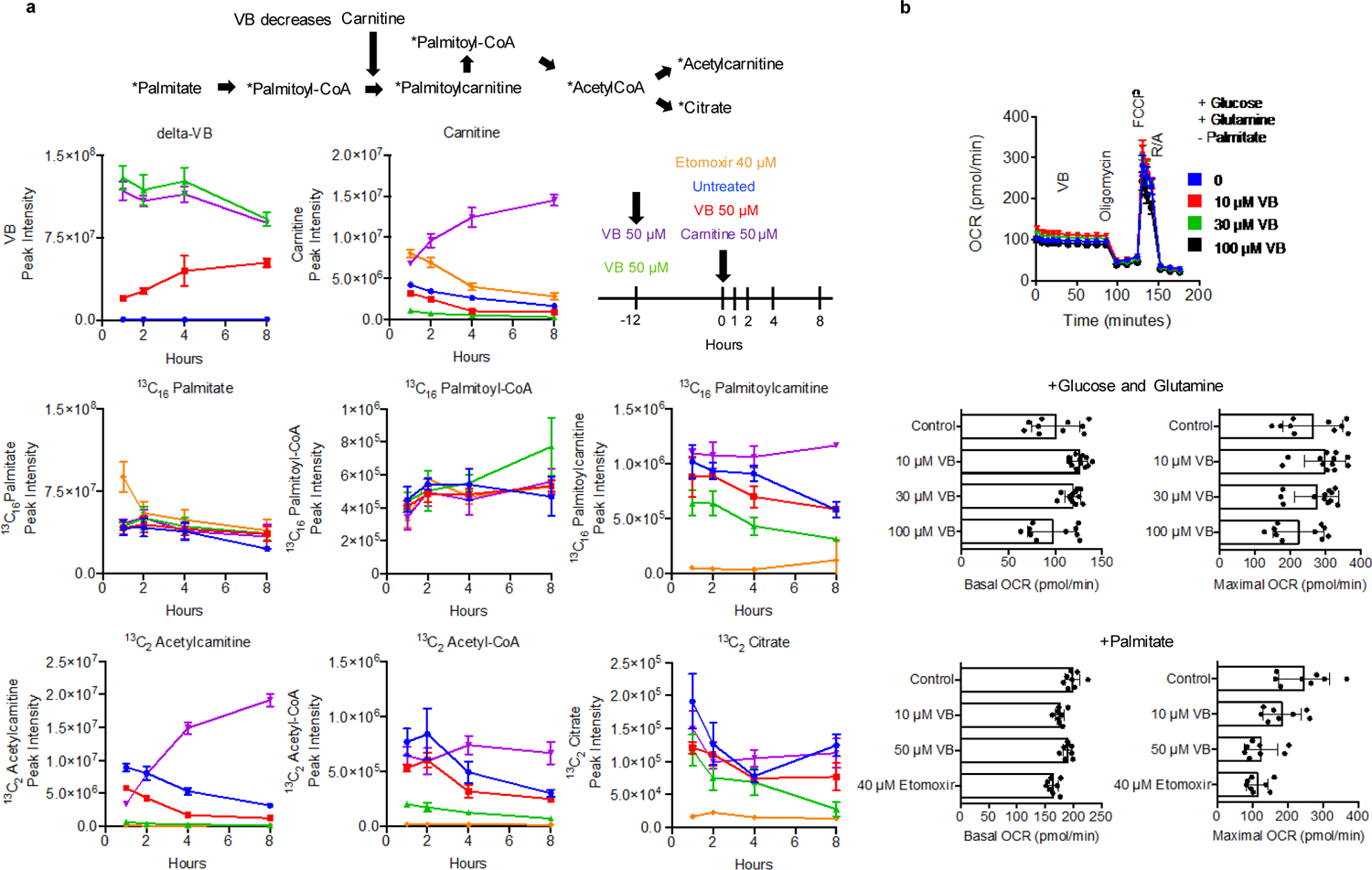
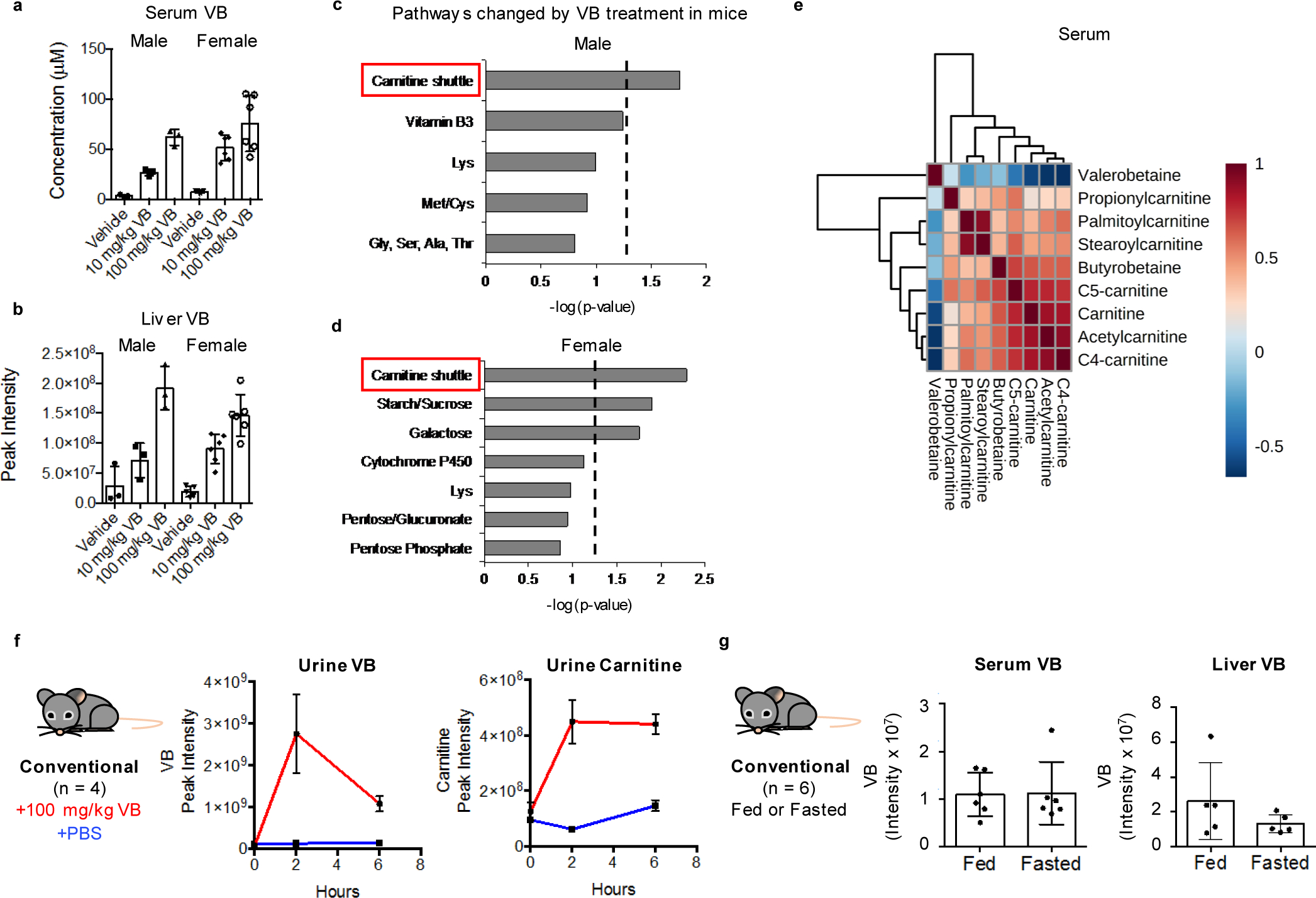
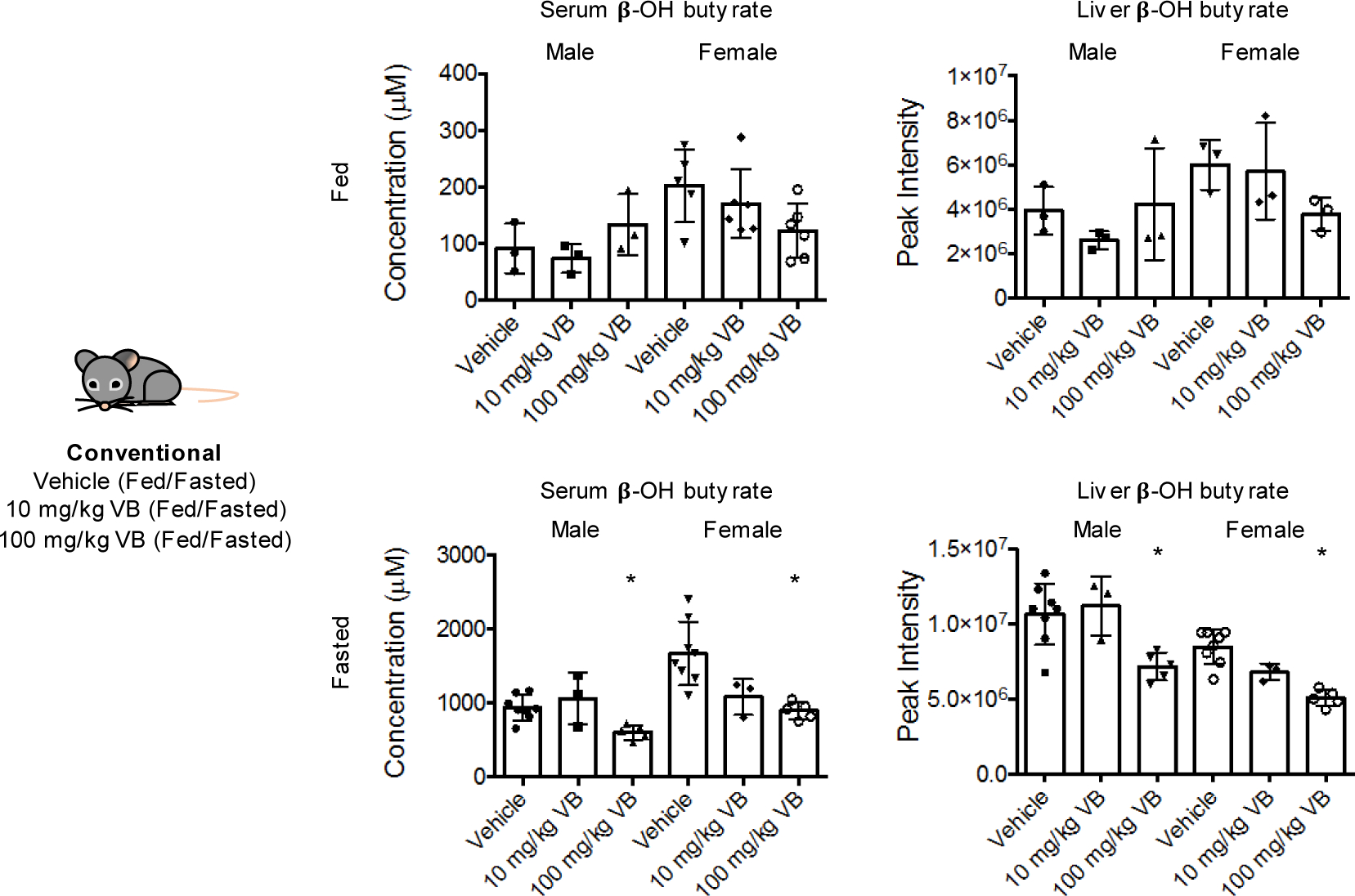
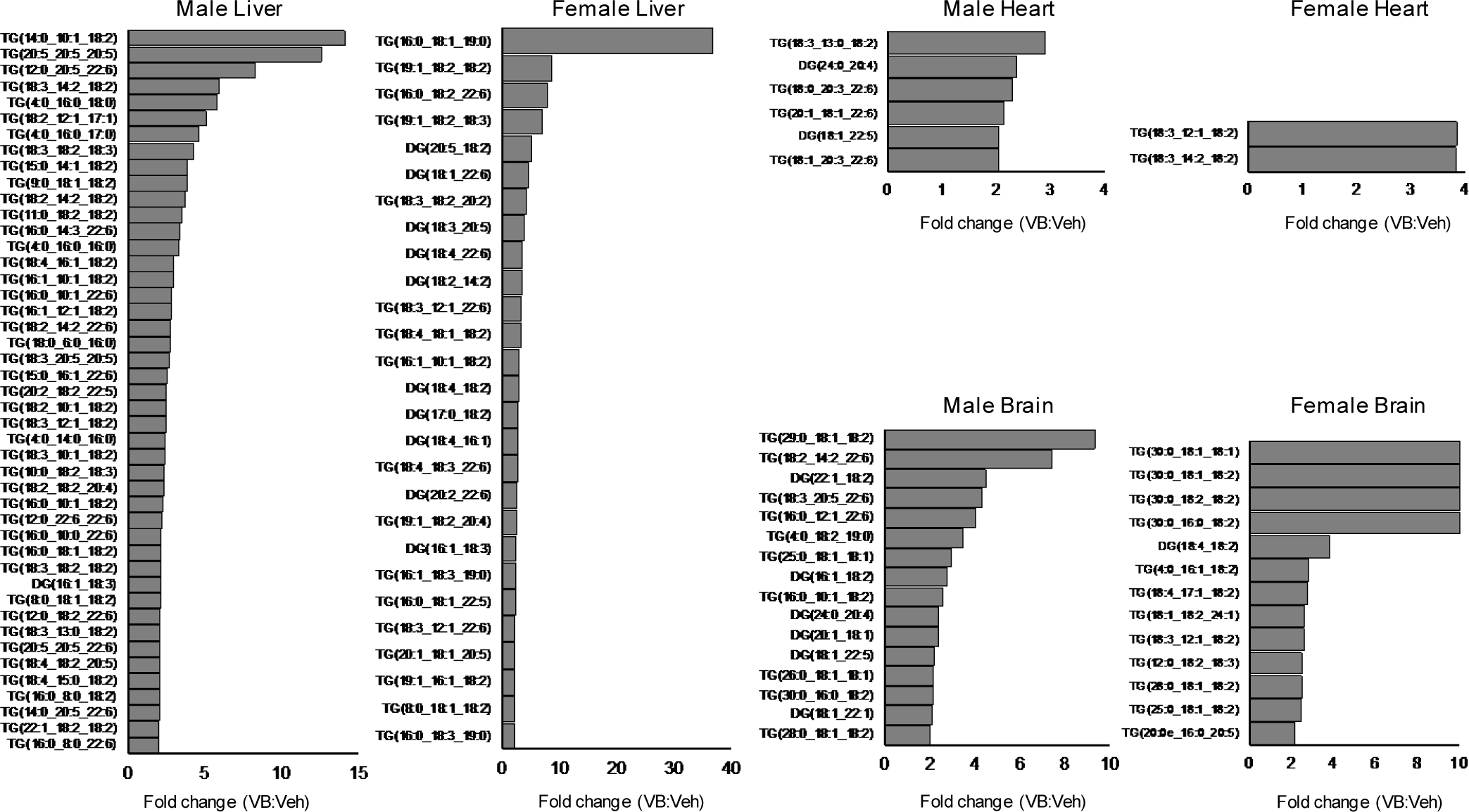
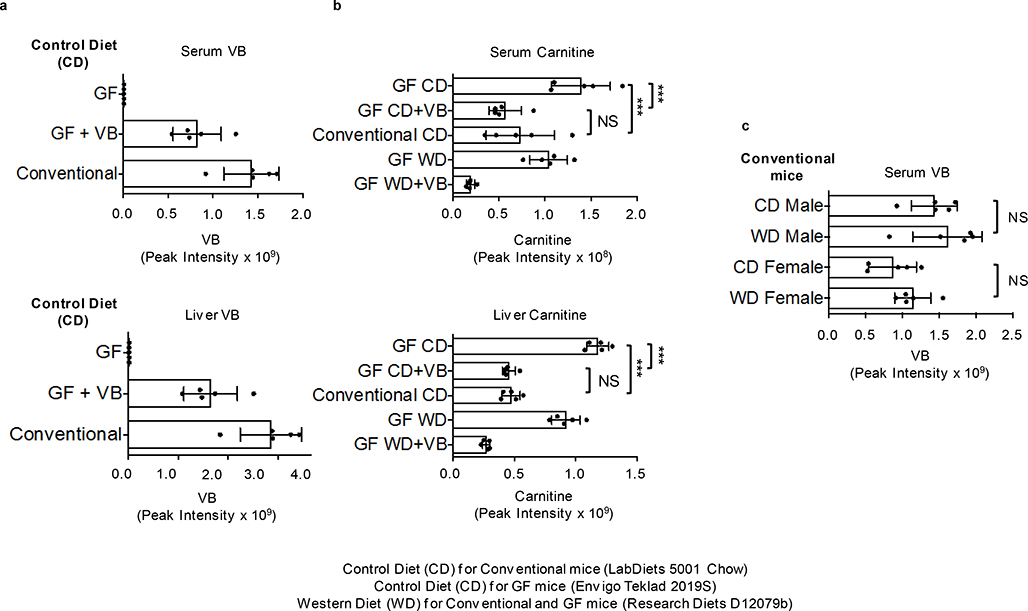
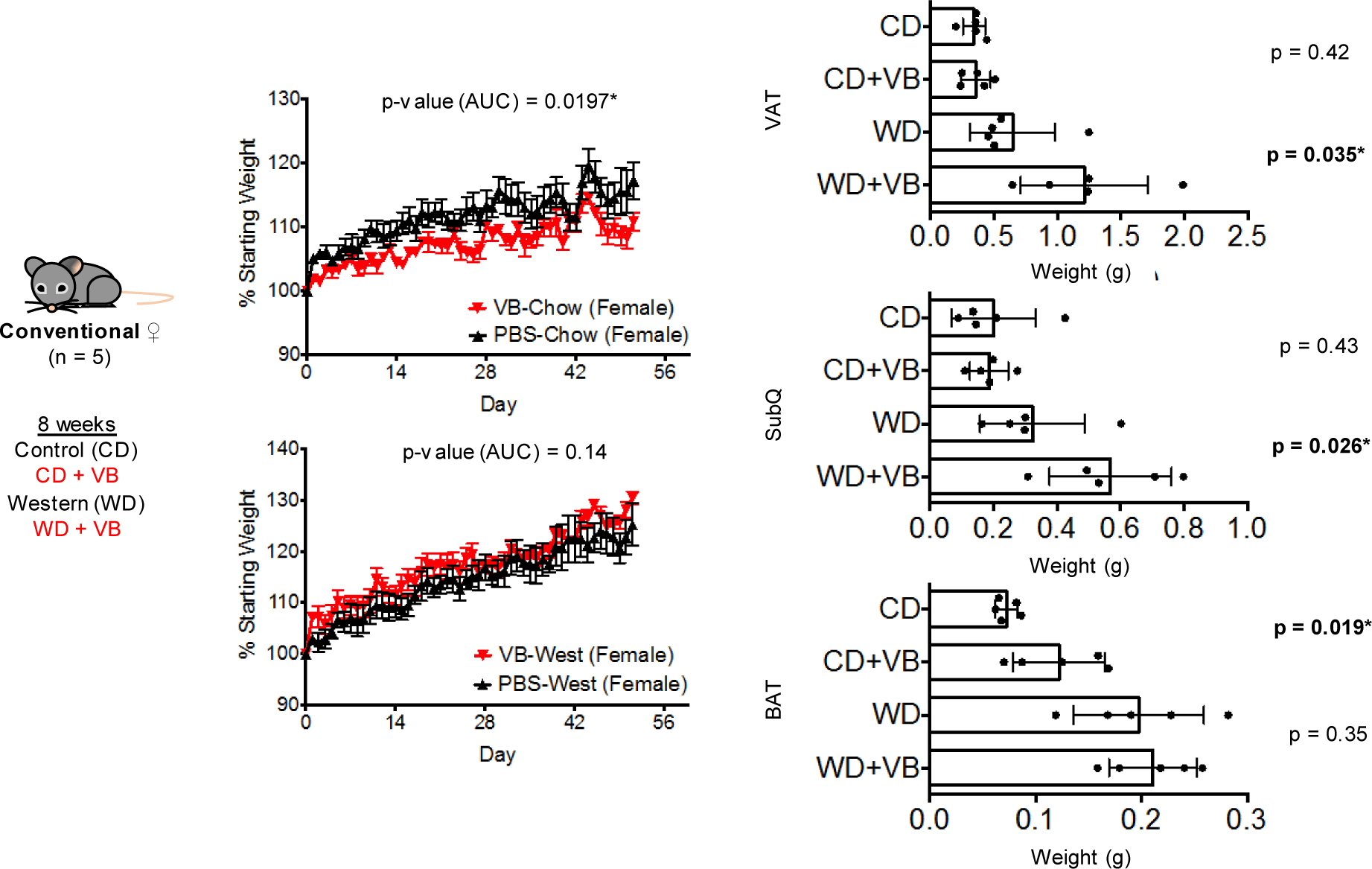
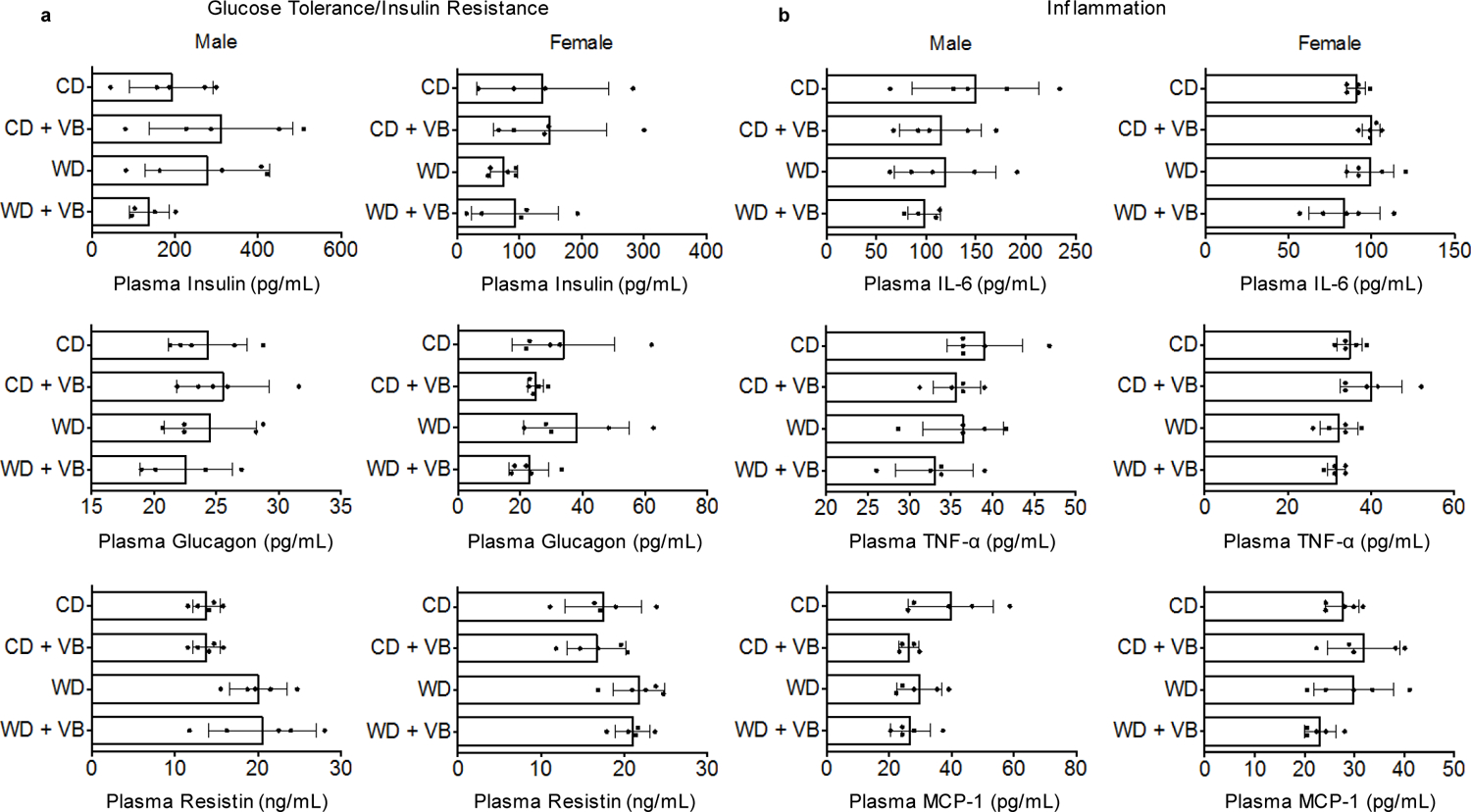
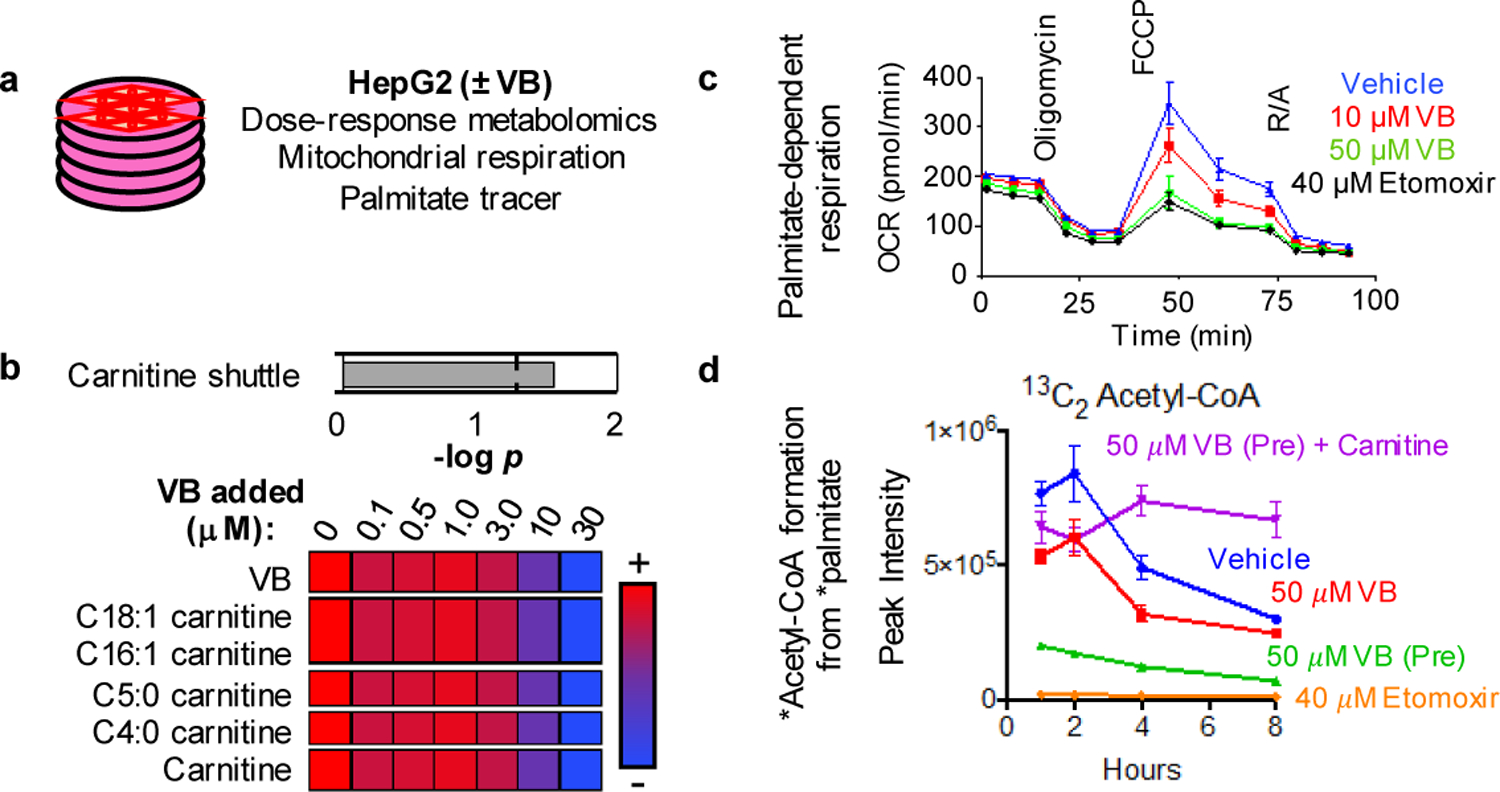
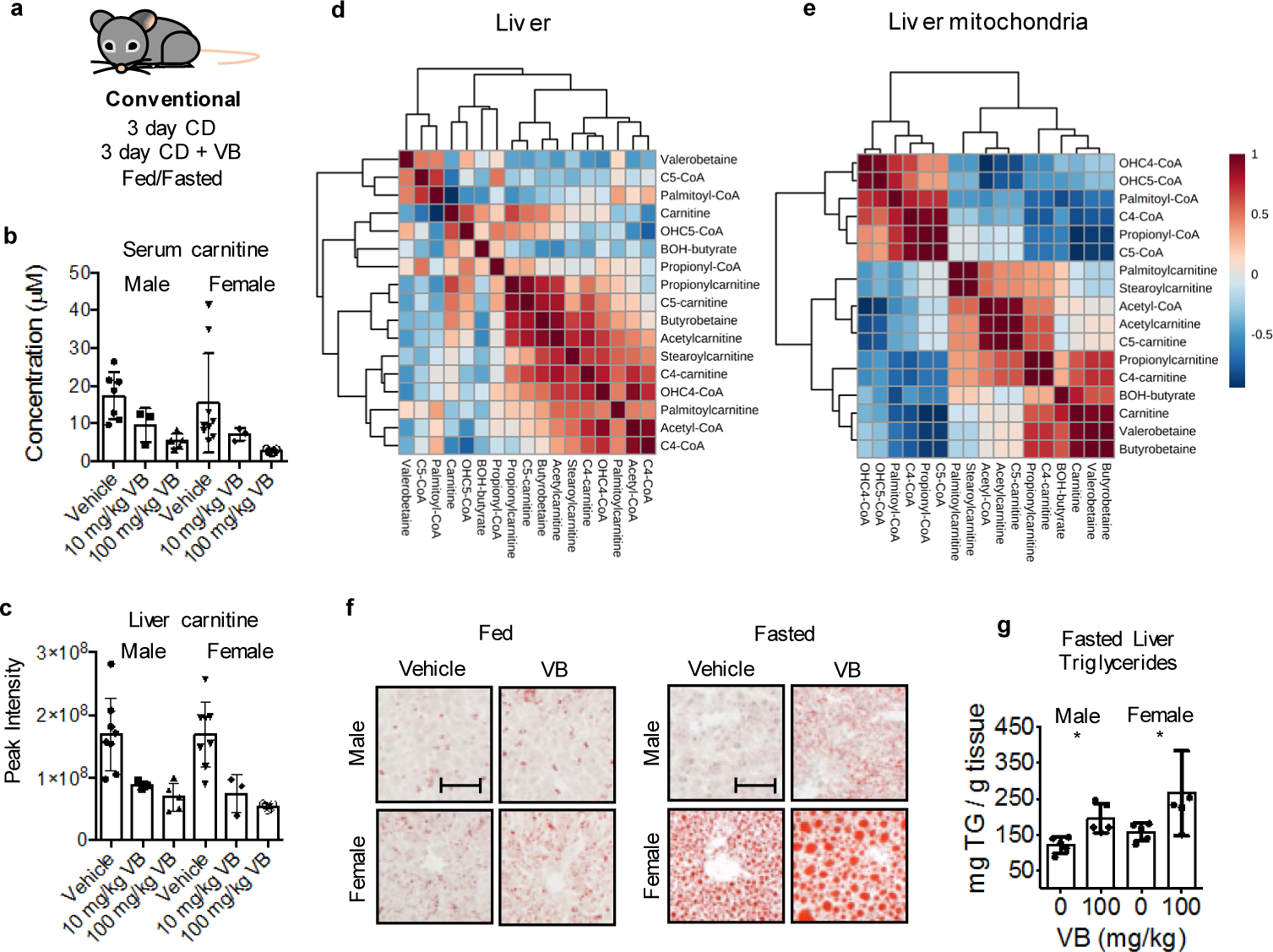
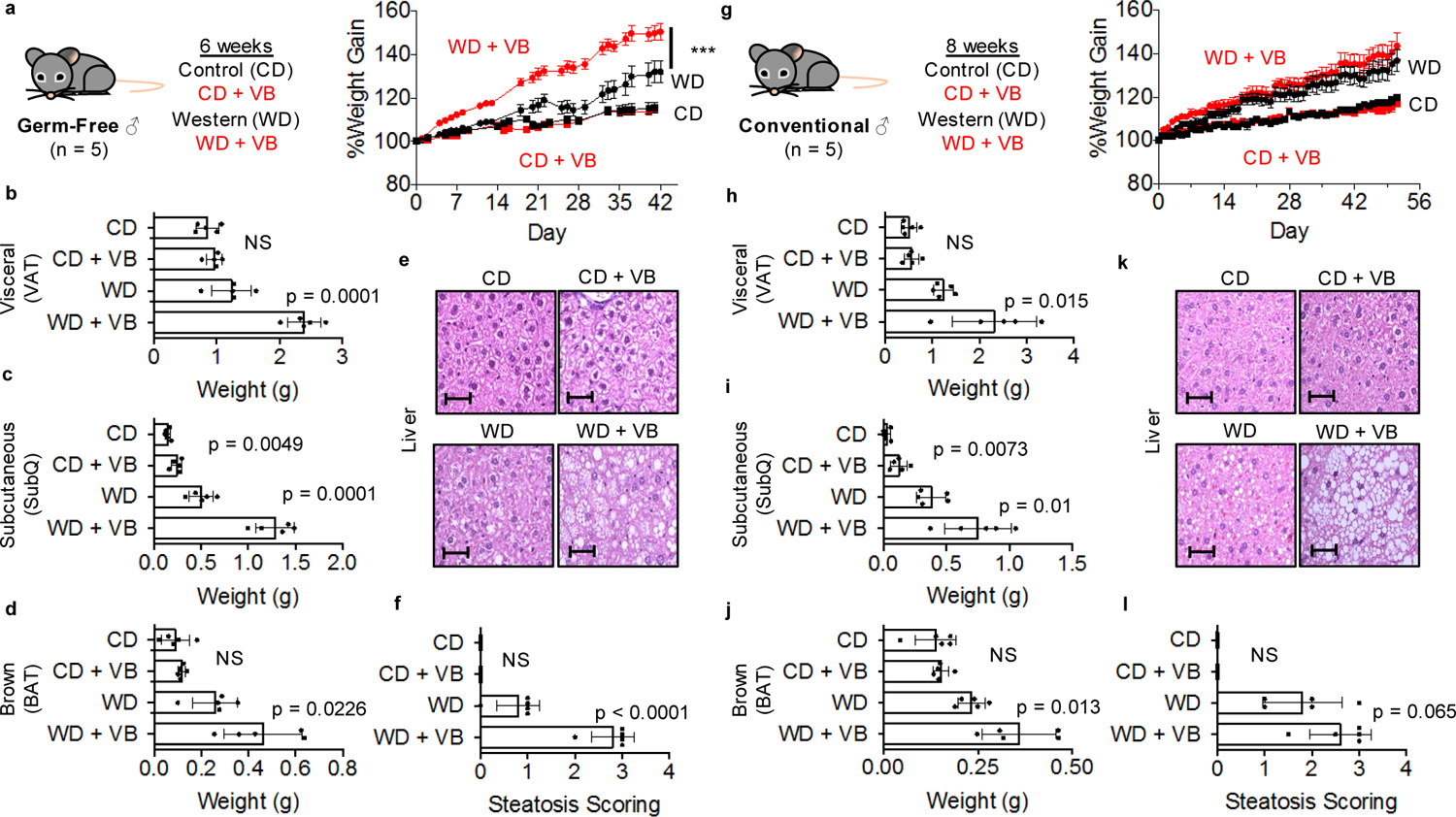
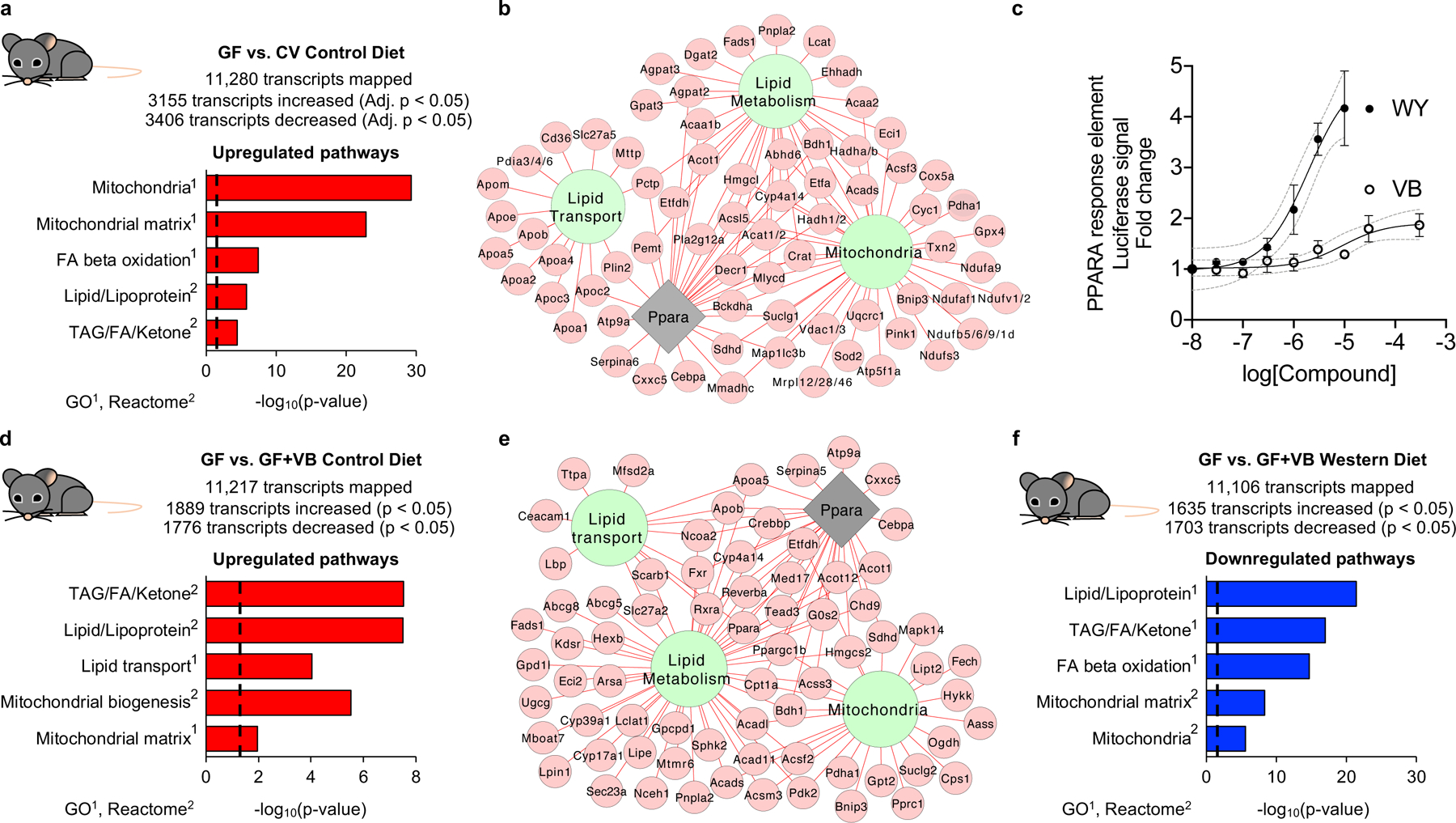
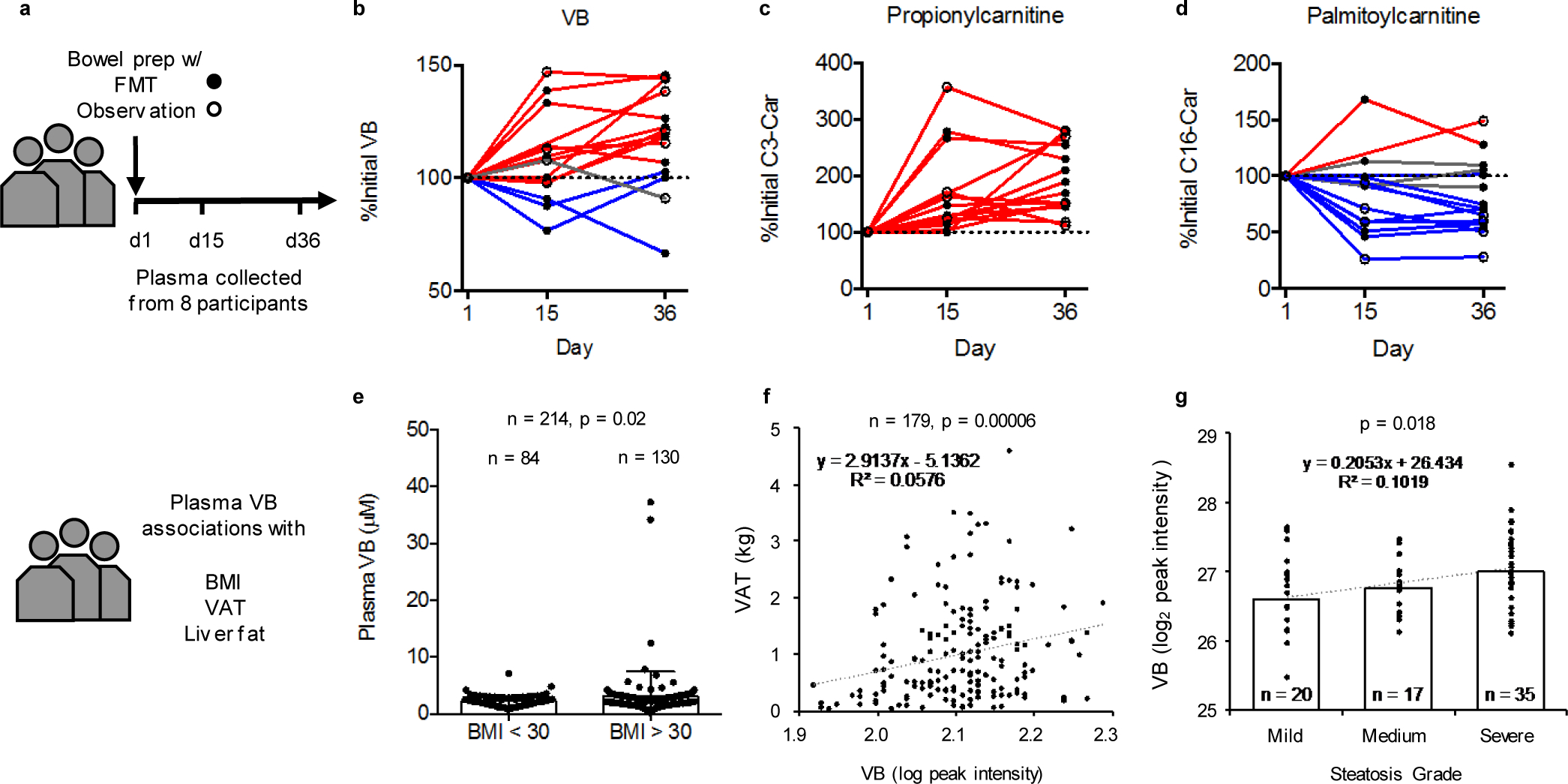
Obesity and obesity-related metabolic disorders are linked to the intestinal microbiome. However, the causality of changes in the microbiome-host interaction affecting energy metabolism remains controversial. Here, we show the microbiome-derived metabolite δ-valerobetaine (VB) is a diet-dependent obesogen that is increased with phenotypic obesity and is correlated with visceral adipose tissue mass in humans. VB is absent in germ-free mice and their mitochondria but present in ex-germ-free conventionalized mice and their mitochondria. Mechanistic studies in vivo and in vitro show VB is produced by diverse bacterial species and inhibits mitochondrial fatty acid oxidation through decreasing cellular carnitine and mitochondrial long-chain acyl-coenzyme As. VB administration to germ-free and conventional mice increases visceral fat mass and exacerbates hepatic steatosis with a western diet but not control diet. Thus, VB provides a molecular target to understand and potentially manage microbiome-host symbiosis or dysbiosis in diet-dependent obesity.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
Authors have no conflict of interest.
Figures
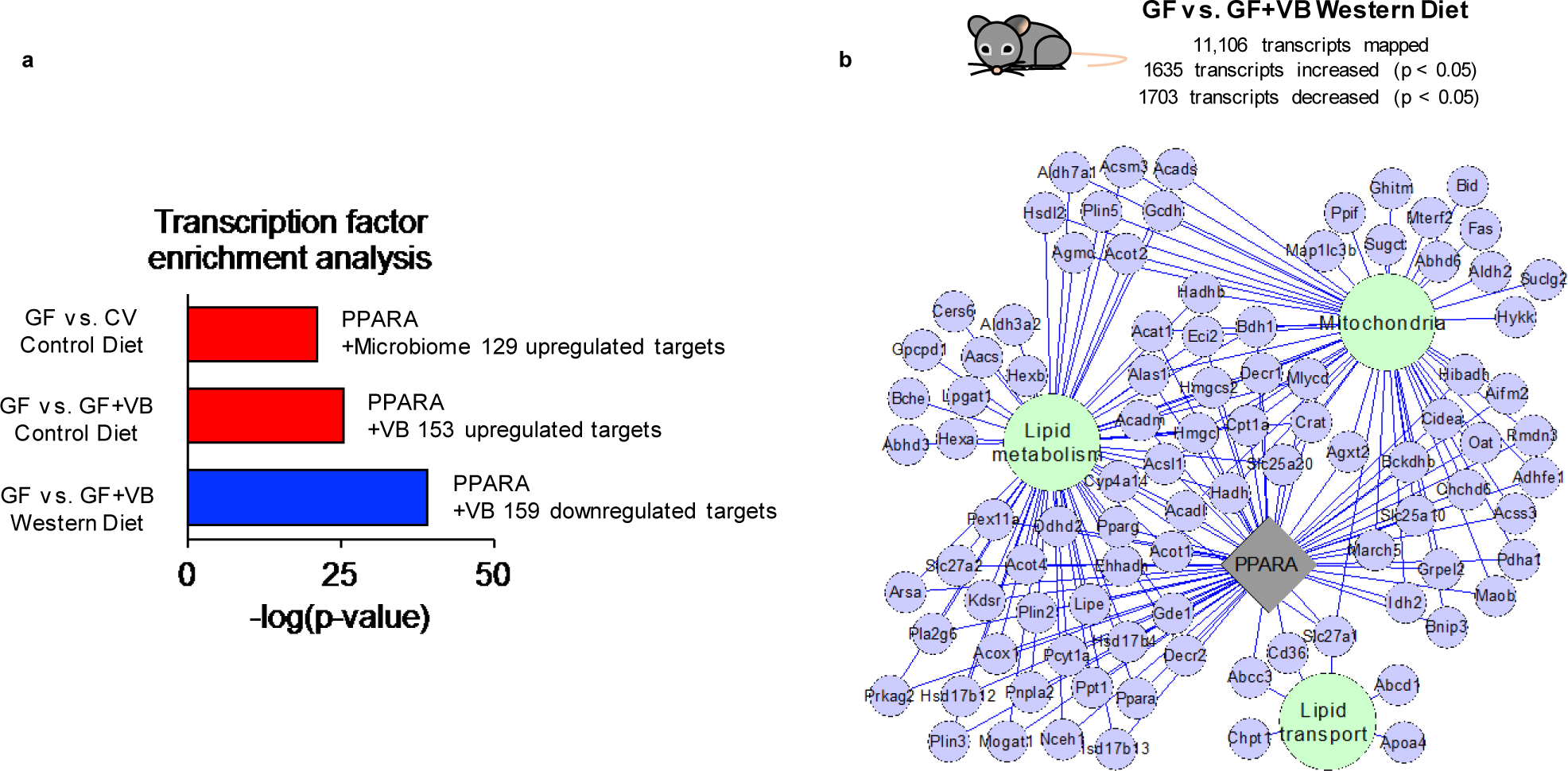
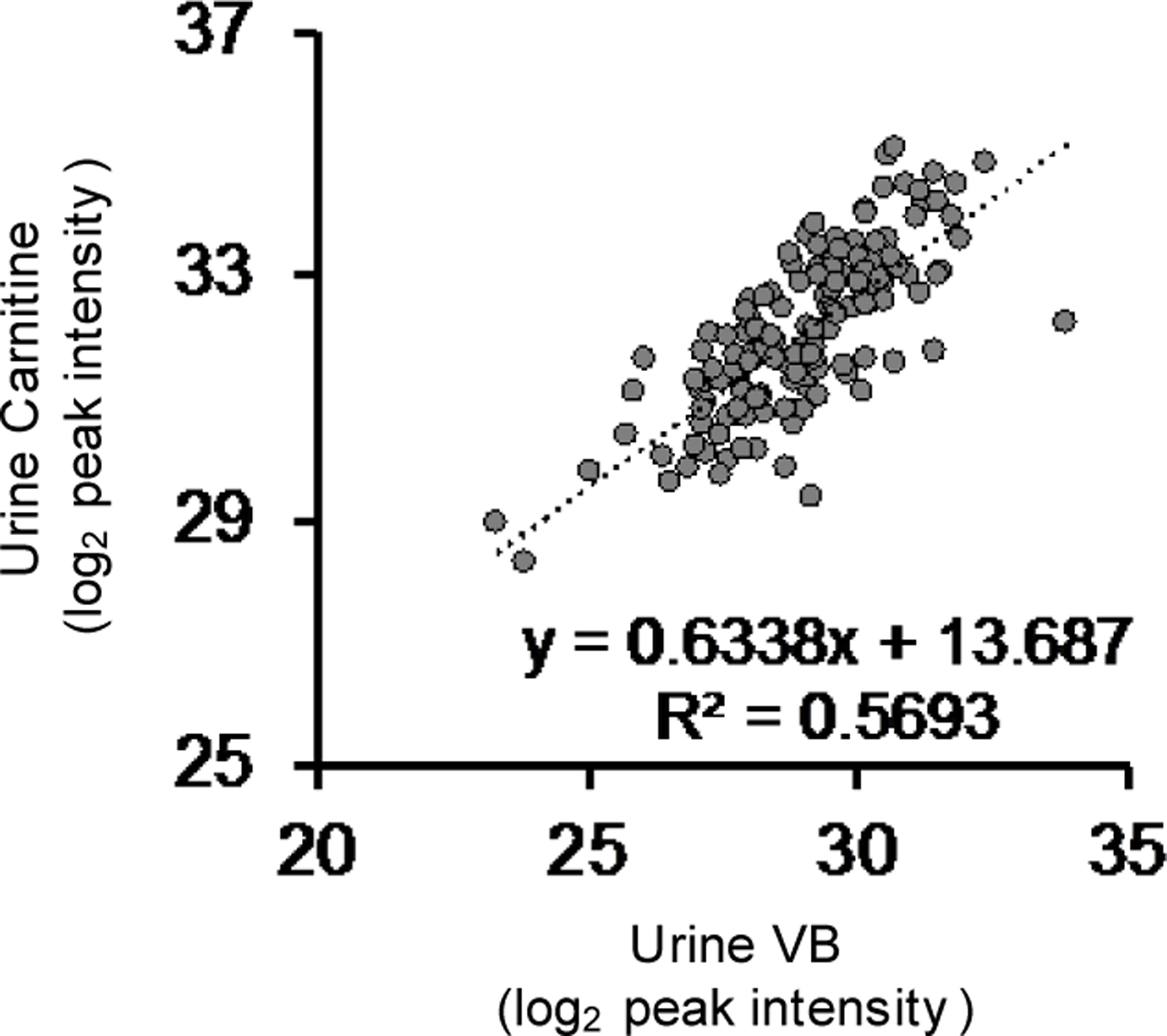
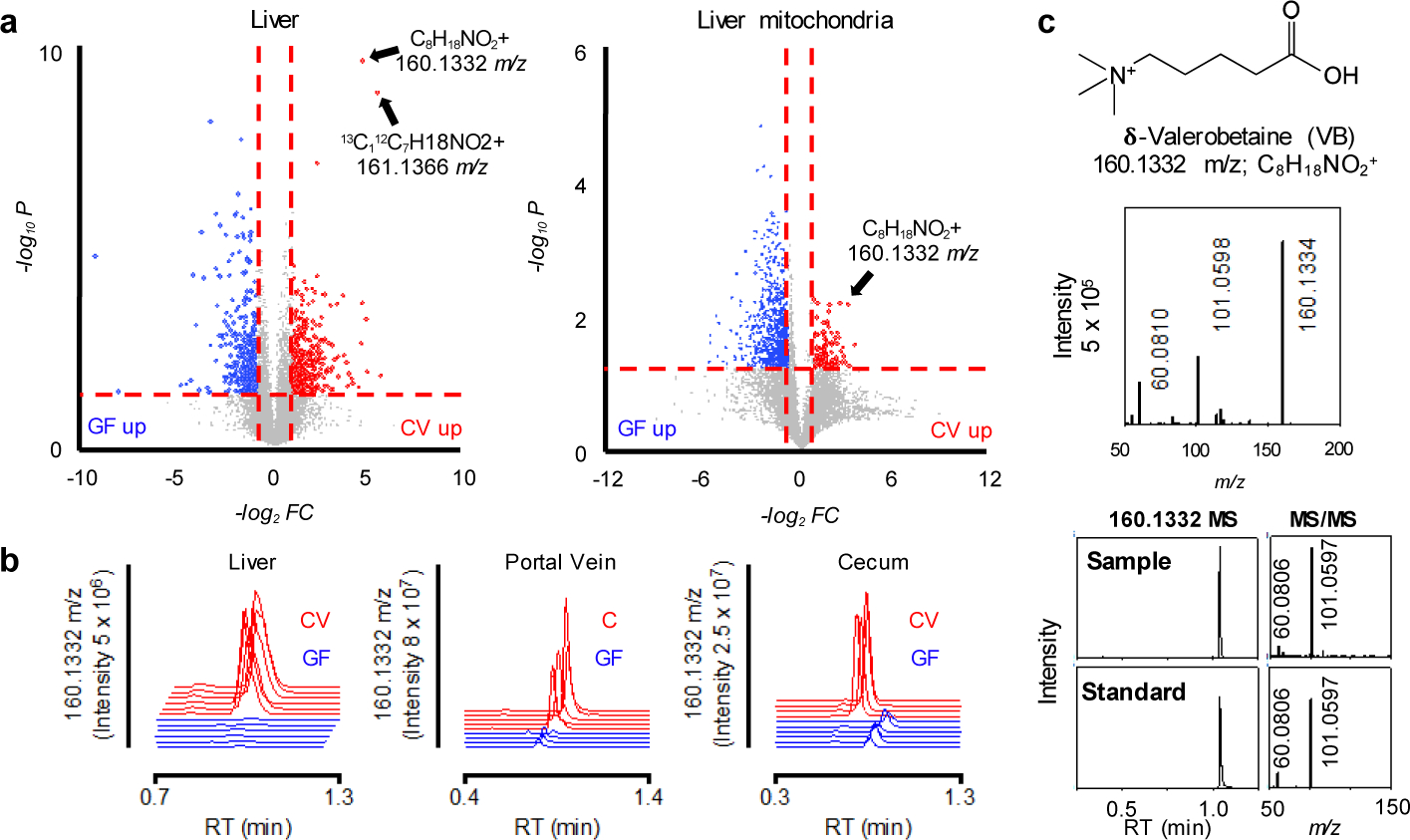
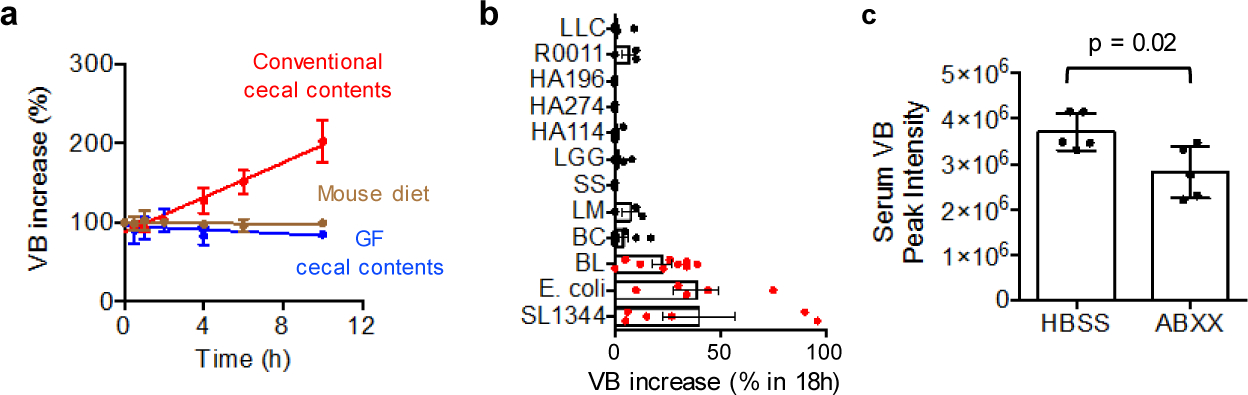
Comment in
-
Inconsistent nomenclature of microbiota-associated metabolites hampers progress of the field.Nat Metab. 2022 Apr;4(4):406. doi: 10.1038/s42255-022-00562-4. Nat Metab. 2022. PMID: 35383295 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
- K24 DK096574/DK/NIDDK NIH HHS/United States
- R00 AA021803/AA/NIAAA NIH HHS/United States
- S10 OD018006/OD/NIH HHS/United States
- R01 AA026086/AA/NIAAA NIH HHS/United States
- R03 AG066559/AG/NIA NIH HHS/United States
- UL1 TR002378/TR/NCATS NIH HHS/United States
- R01 DK125701/DK/NIDDK NIH HHS/United States
- R01 ES023485/ES/NIEHS NIH HHS/United States
- P30 ES019776/ES/NIEHS NIH HHS/United States
- K01 DK102851/DK/NIDDK NIH HHS/United States
- R03 DK117246/DK/NIDDK NIH HHS/United States
- U2C ES030163/ES/NIEHS NIH HHS/United States
- R21 HD089056/HD/NICHD NIH HHS/United States
- UH2 AI132345/AI/NIAID NIH HHS/United States
- T32 GM008602/GM/NIGMS NIH HHS/United States
- RC2 DK118619/DK/NIDDK NIH HHS/United States
- R21 ES031824/ES/NIEHS NIH HHS/United States
- R01 AI064462/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
