

Tim-3 regulates sepsis-induced immunosuppression by inhibiting the NF-κB signaling pathway in CD4 T cells
- PMID: 34933101
- PMCID: PMC8899604
- DOI: 10.1016/j.ymthe.2021.12.013
Tim-3 regulates sepsis-induced immunosuppression by inhibiting the NF-κB signaling pathway in CD4 T cells
Abstract
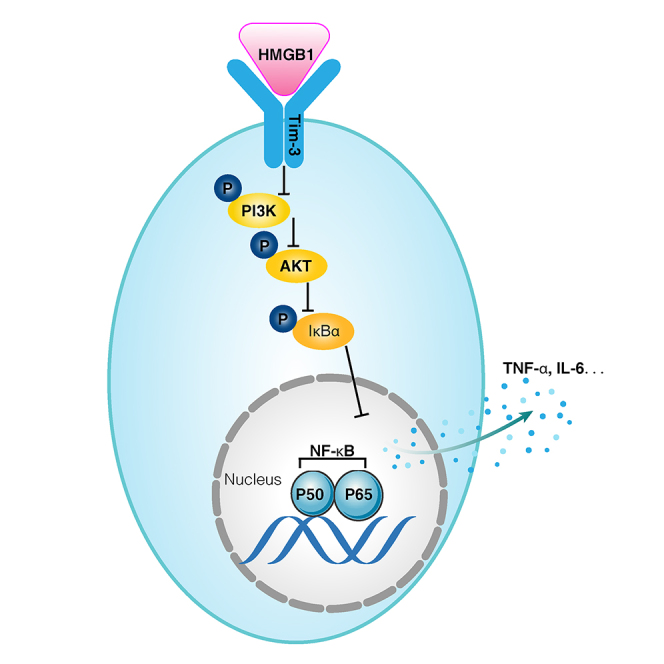
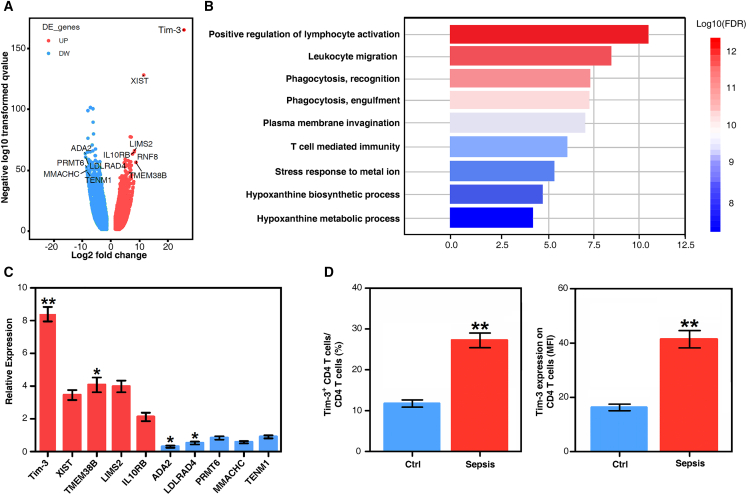
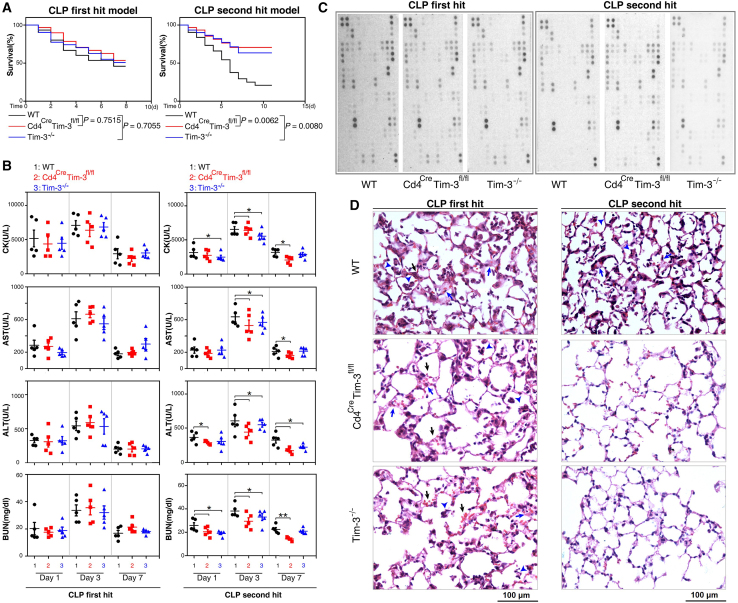
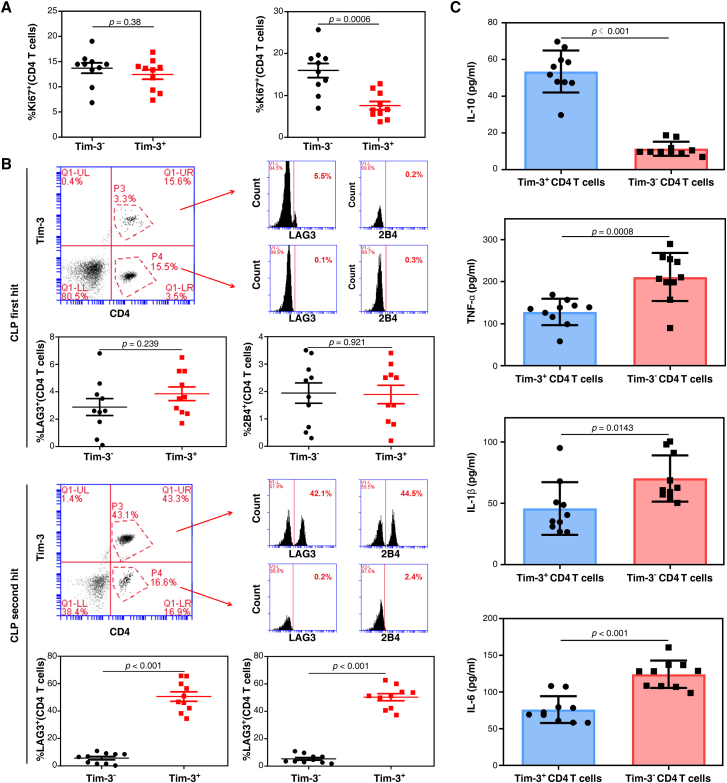
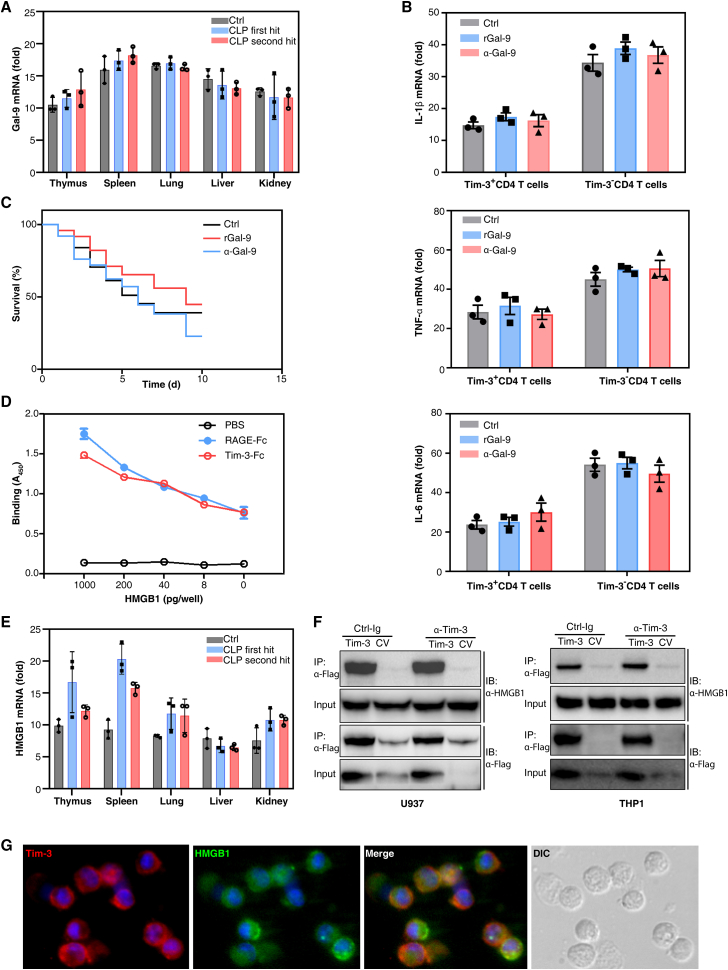
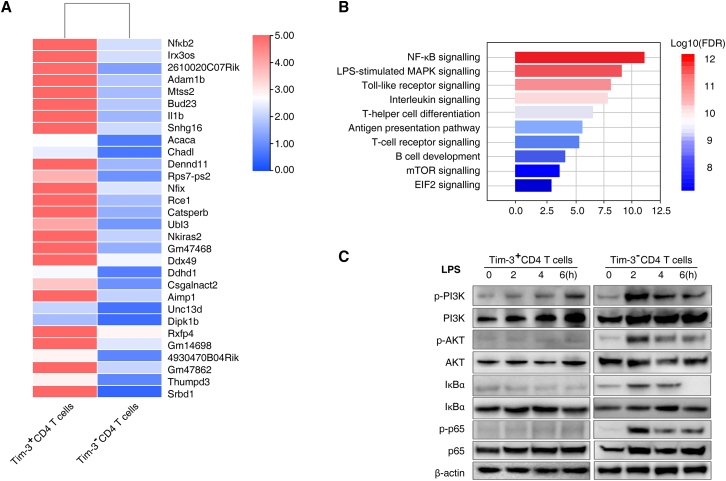
Immunosuppression in response to severe sepsis remains a serious human health concern. Evidence of sepsis-induced immunosuppression includes impaired T lymphocyte function, T lymphocyte depletion or exhaustion, increased susceptibility to opportunistic nosocomial infection, and imbalanced cytokine secretion. CD4 T cells play a critical role in cellular and humoral immune responses during sepsis. Here, using an RNA sequencing assay, we found that the expression of T cell-containing immunoglobulin and mucin domain-3 (Tim-3) on CD4 T cells in sepsis-induced immunosuppression patients was significantly elevated. Furthermore, the percentage of Tim-3+ CD4 T cells from sepsis patients was correlated with the mortality of sepsis-induced immunosuppression. Conditional deletion of Tim-3 in CD4 T cells and systemic Tim-3 deletion both reduced mortality in response to sepsis in mice by preserving organ function. Tim-3+ CD4 T cells exhibited reduced proliferative ability and elevated expression of inhibitory markers compared with Tim-3-CD4 T cells. Colocalization analyses indicated that HMGB1 was a ligand that binds to Tim-3 on CD4 T cells and that its binding inhibited the NF-κB signaling pathway in Tim-3+ CD4 T cells during sepsis-induced immunosuppression. Together, our findings reveal the mechanism of Tim-3 in regulating sepsis-induced immunosuppression and provide a novel therapeutic target for this condition.
Keywords: CD4 T cells; T cell-containing immunoglobulin and mucin domain-3 (Tim-3); biomarker; sepsis-induced immunosuppression.
Copyright © 2021 The American Society of Gene and Cell Therapy. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests All authors declare that they have no competing financial interests.
Figures
References
-
- Van Der Poll T., Van De Veerdonk F.L., Scicluna B.P., Netea M.G. The immunopathology of sepsis and potential therapeutic targets. Nat. Rev. Immunol. 2017;17:407–420. - PubMed
-
- Fattahi F., Ward P.A. Understanding immunosuppression after sepsis. Immunity. 2017;47:3–5. - PubMed
-
- Gotts J.E., Matthay M.A. Sepsis: pathophysiology and clinical management. BMJ. 2016;353:i1585. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
