

Local and systemic responses to SARS-CoV-2 infection in children and adults
- PMID: 34937051
- PMCID: PMC8828466
- DOI: 10.1038/s41586-021-04345-x
Local and systemic responses to SARS-CoV-2 infection in children and adults
Abstract
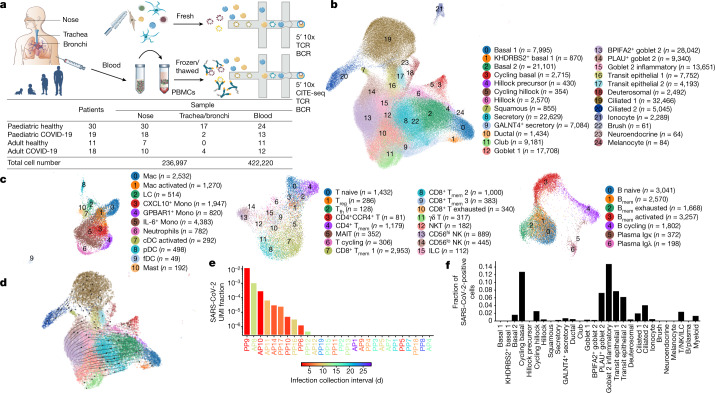
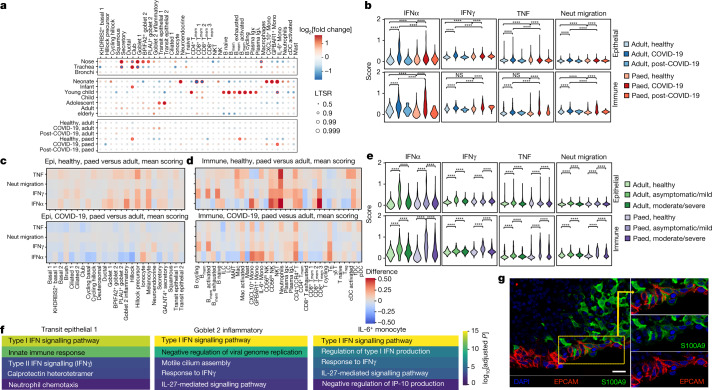
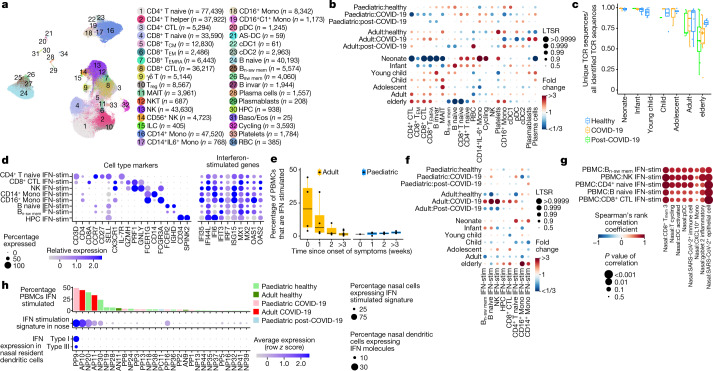
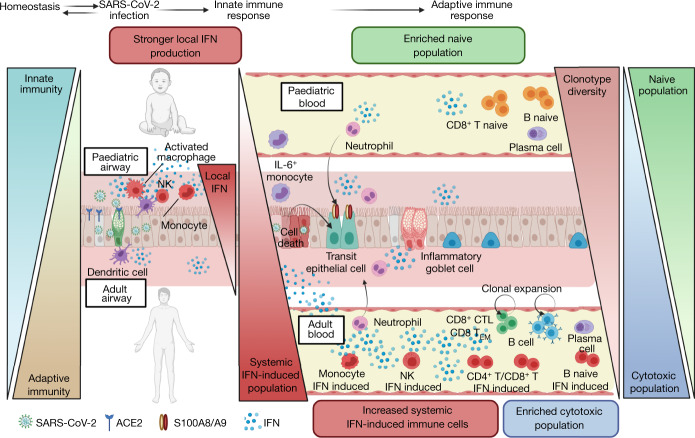
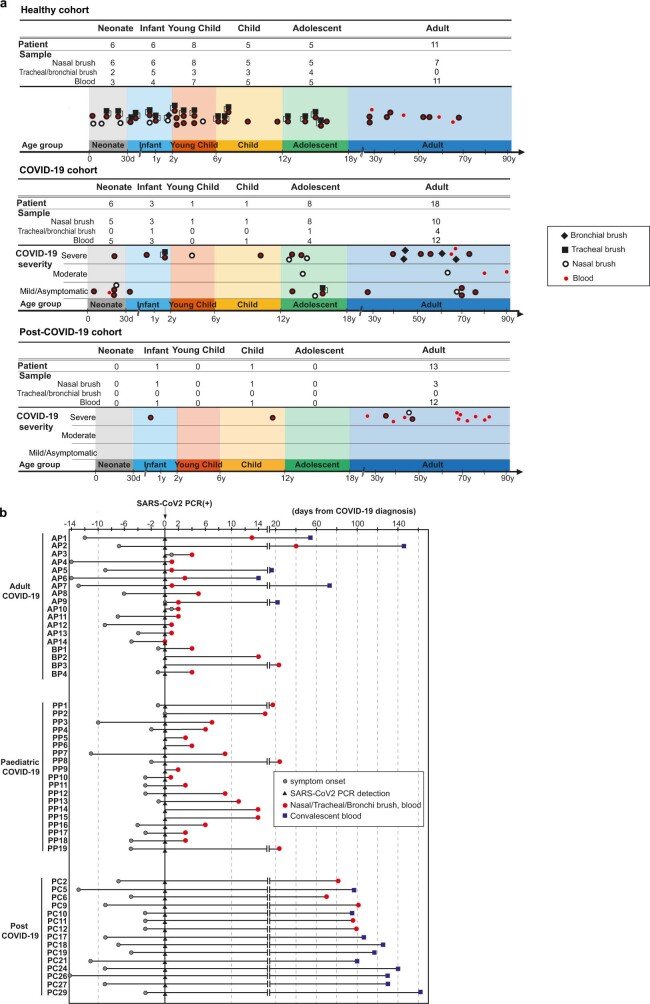
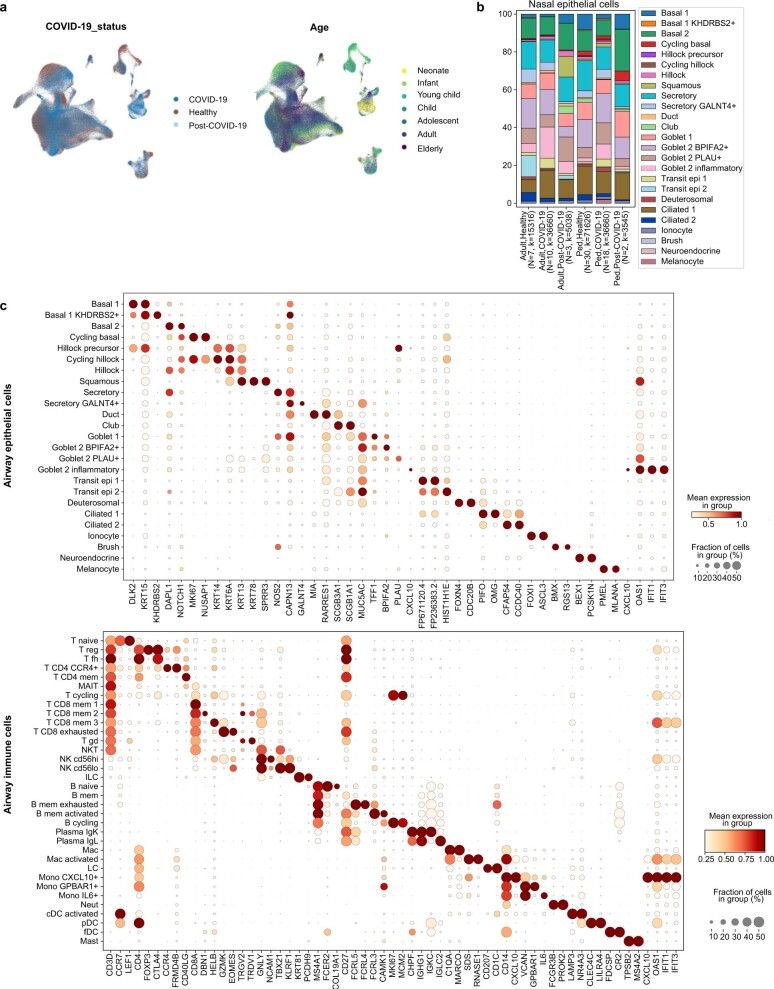
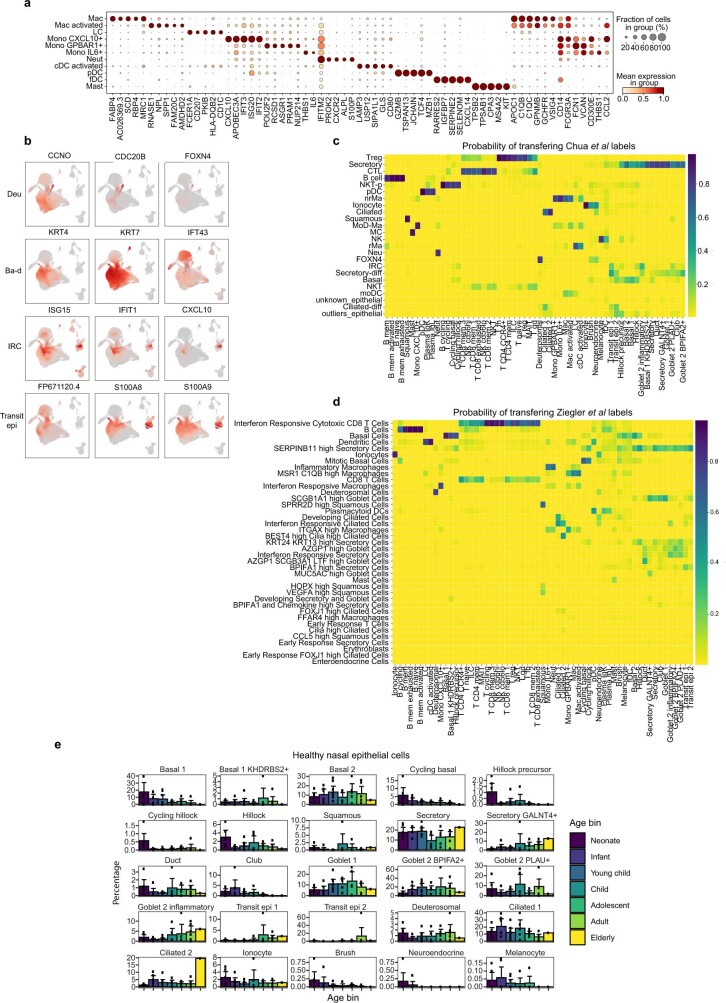
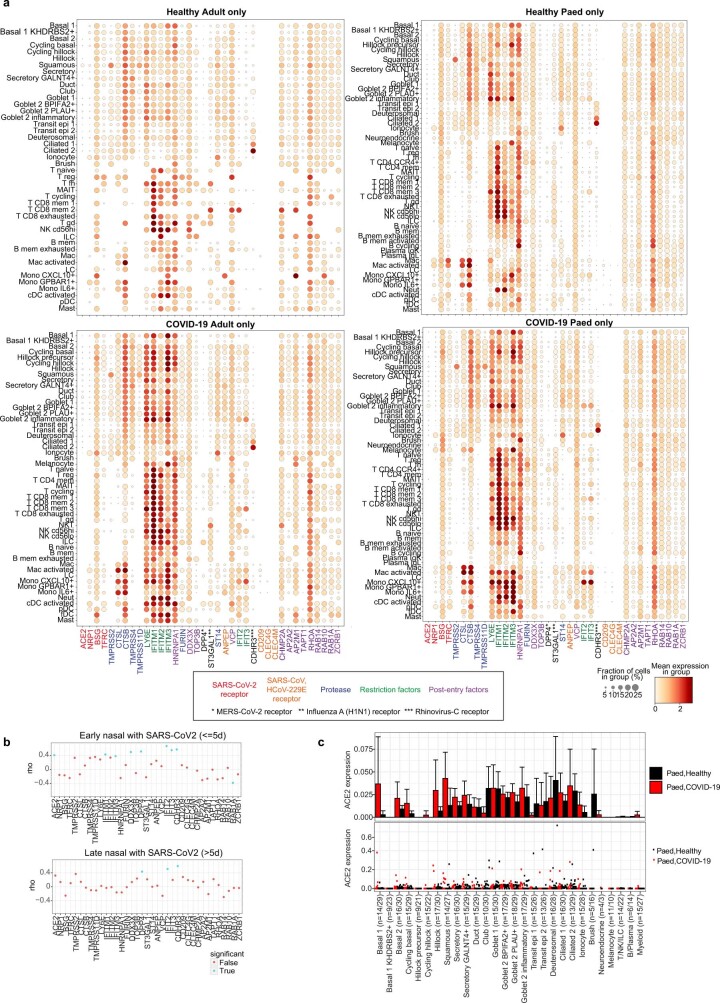
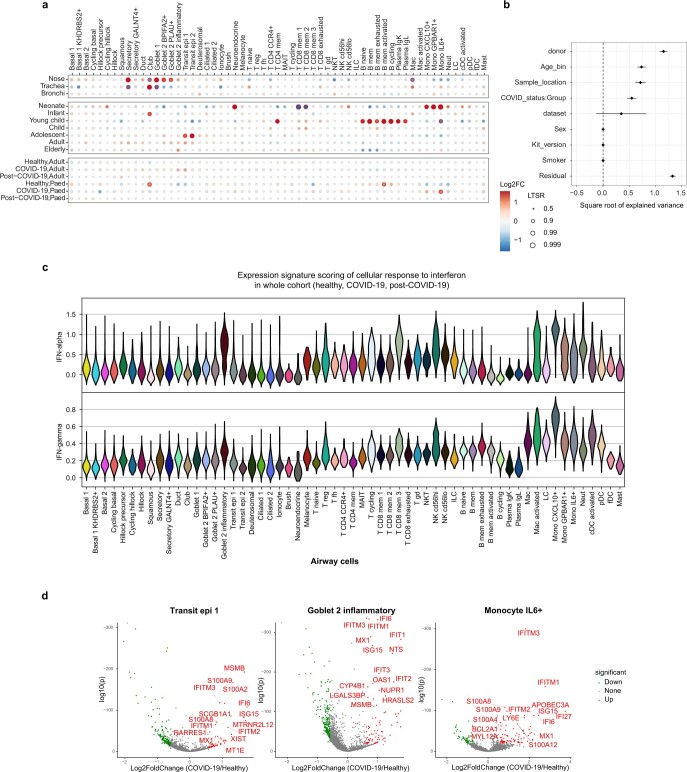
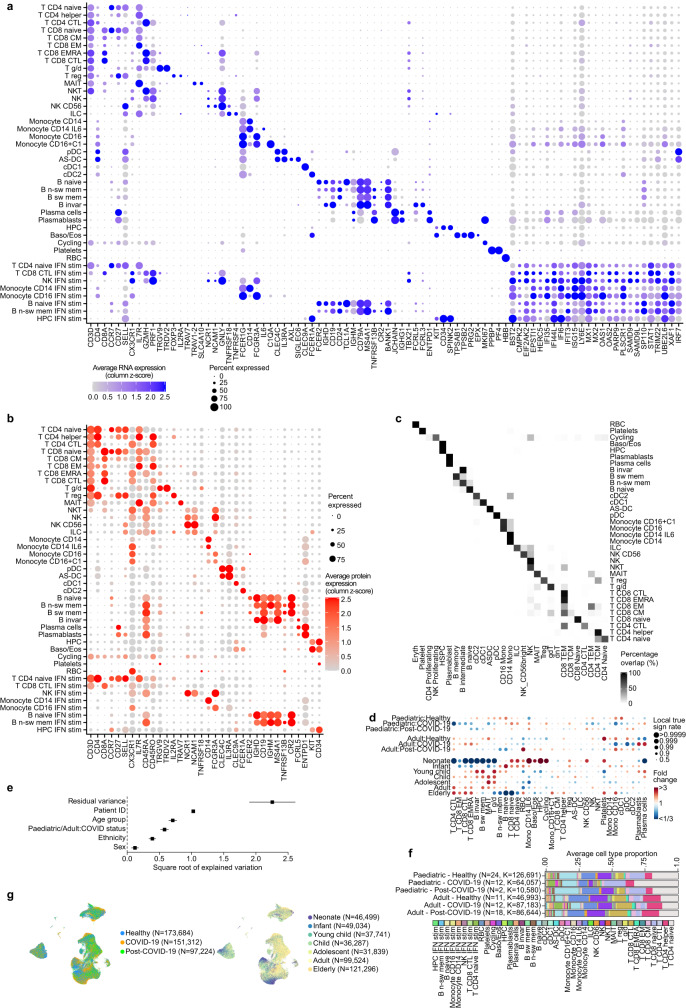
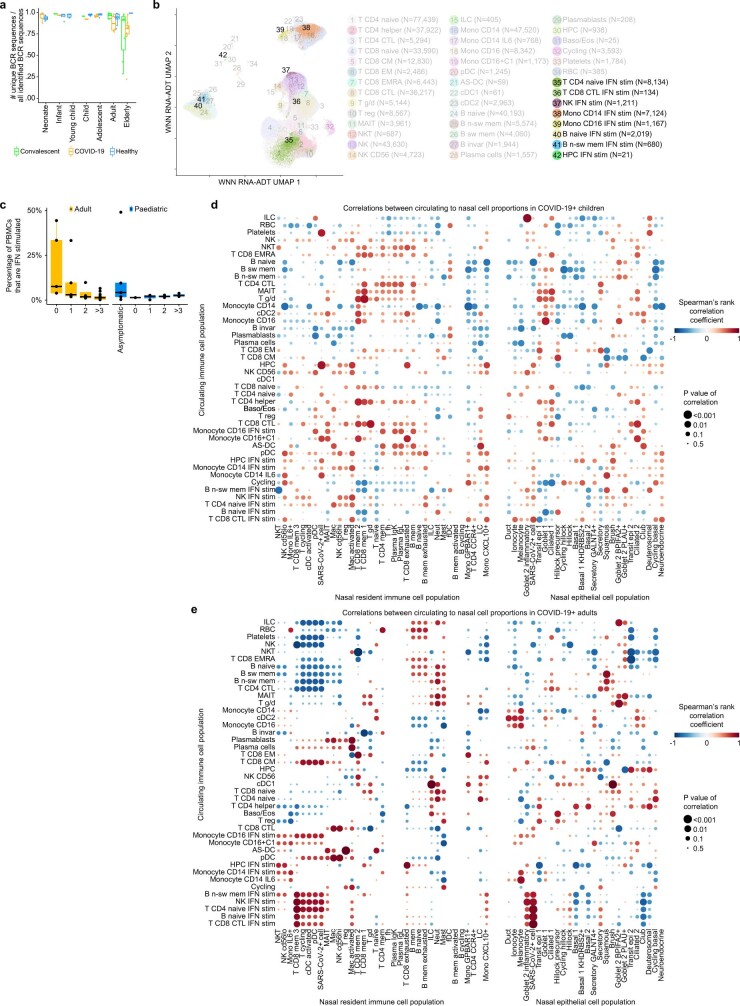
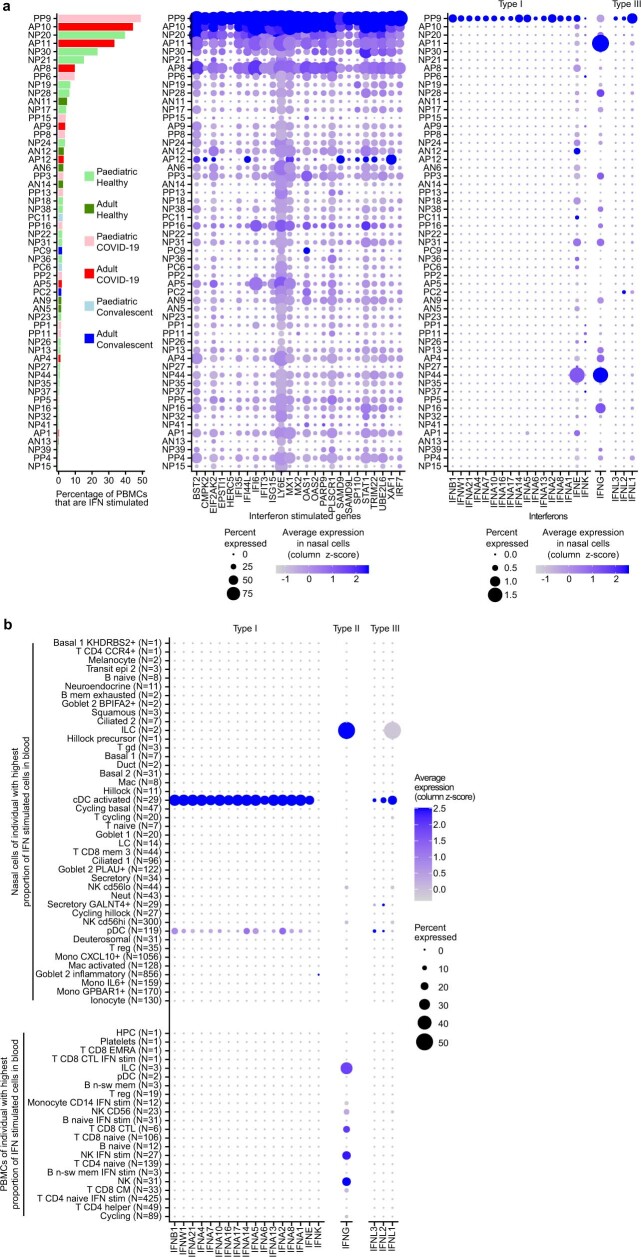
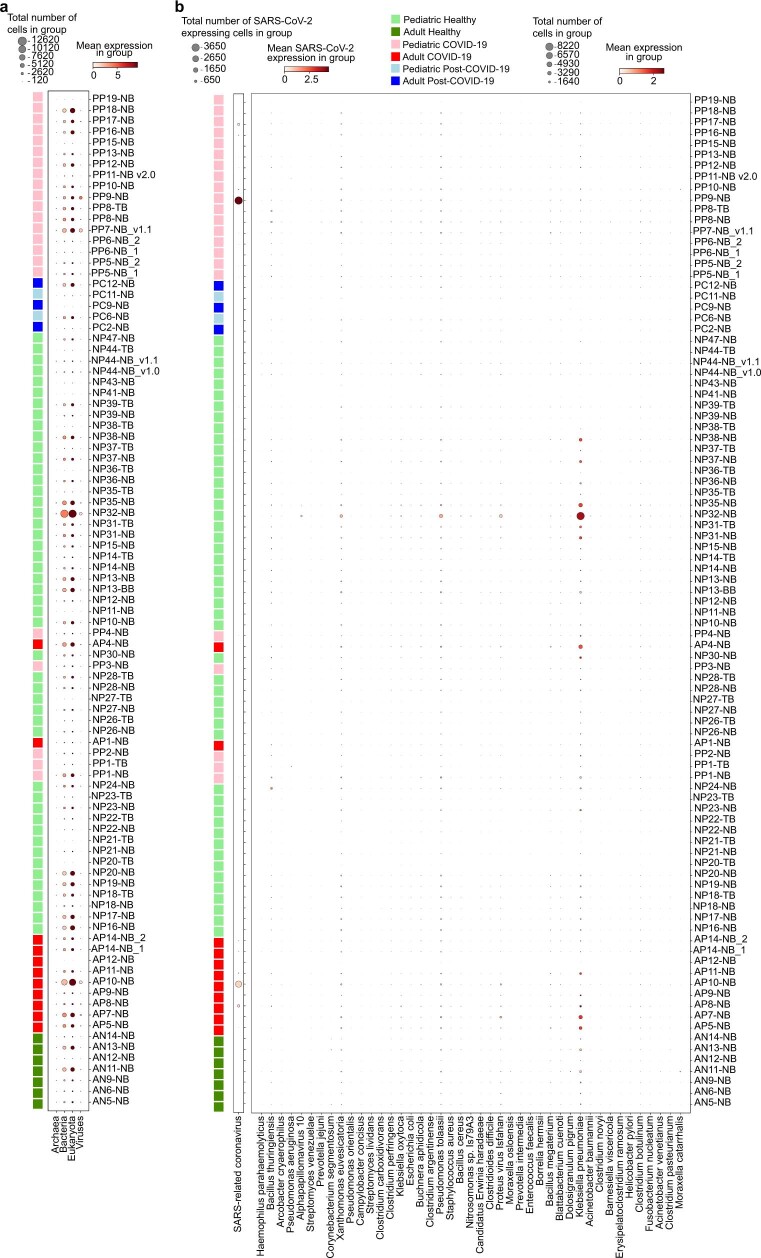
It is not fully understood why COVID-19 is typically milder in children1-3. Here, to examine the differences between children and adults in their response to SARS-CoV-2 infection, we analysed paediatric and adult patients with COVID-19 as well as healthy control individuals (total n = 93) using single-cell multi-omic profiling of matched nasal, tracheal, bronchial and blood samples. In the airways of healthy paediatric individuals, we observed cells that were already in an interferon-activated state, which after SARS-CoV-2 infection was further induced especially in airway immune cells. We postulate that higher paediatric innate interferon responses restrict viral replication and disease progression. The systemic response in children was characterized by increases in naive lymphocytes and a depletion of natural killer cells, whereas, in adults, cytotoxic T cells and interferon-stimulated subpopulations were significantly increased. We provide evidence that dendritic cells initiate interferon signalling in early infection, and identify epithelial cell states associated with COVID-19 and age. Our matching nasal and blood data show a strong interferon response in the airways with the induction of systemic interferon-stimulated populations, which were substantially reduced in paediatric patients. Together, we provide several mechanisms that explain the milder clinical syndrome observed in children.
© 2021. The Author(s).
Conflict of interest statement
In the past three years, S.A.T. has worked as a consultant for Genentech, Roche and Transition Bio, and is a remunerated member of the Scientific Advisory Boards of Qiagen, GlaxoSmithKline and Foresite Labs and an equity holder of Transition Bio. P.M. is a Medical Research Council-GlaxoSmithKline (MRC-GSK) Experimental Medicine Initiative to Explore New Therapies (EMINENT) clinical training fellow with project funding, has served on an advisory board for SOBI, outside the submitted work, and receives co-funding by the National Institute for Health Research (NIHR) University College London Hospitals Biomedical Research Centre (UCLH BRC).
Figures
References
-
- Ledford H. Deaths from COVID ‘incredibly rare’ among children. Nature. 2021;595:639–639. doi: 10.1038/d41586-021-01897-w. - DOI
Publication types
MeSH terms
Substances
Grants and funding
- MC_PC_17230/MRC_/Medical Research Council/United Kingdom
- R01 HL153122/HL/NHLBI NIH HHS/United States
- MC_PC_20022/MRC_/Medical Research Council/United Kingdom
- WT211276/Z/18/Z/WT_/Wellcome Trust/United Kingdom
- MR/S005579/1/MRC_/Medical Research Council/United Kingdom
- P01 AG049665/AG/NIA NIH HHS/United States
- P01 HL154998/HL/NHLBI NIH HHS/United States
- MR/K017047/1/MRC_/Medical Research Council/United Kingdom
- R01 HL149883/HL/NHLBI NIH HHS/United States
- MR/W014556/1/MRC_/Medical Research Council/United Kingdom
- MR/R015635/1/MRC_/Medical Research Council/United Kingdom
- T32 HL076139/HL/NHLBI NIH HHS/United States
- U19 AI135964/AI/NIAID NIH HHS/United States
- MR/S036113/1/MRC_/Medical Research Council/United Kingdom
- R01 HL153312/HL/NHLBI NIH HHS/United States
- UL1 TR001422/TR/NCATS NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- MR/S035842/1/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
