

Temperature-responsive optogenetic probes of cell signaling
- PMID: 34937907
- PMCID: PMC9252025
- DOI: 10.1038/s41589-021-00917-0
Temperature-responsive optogenetic probes of cell signaling
Abstract
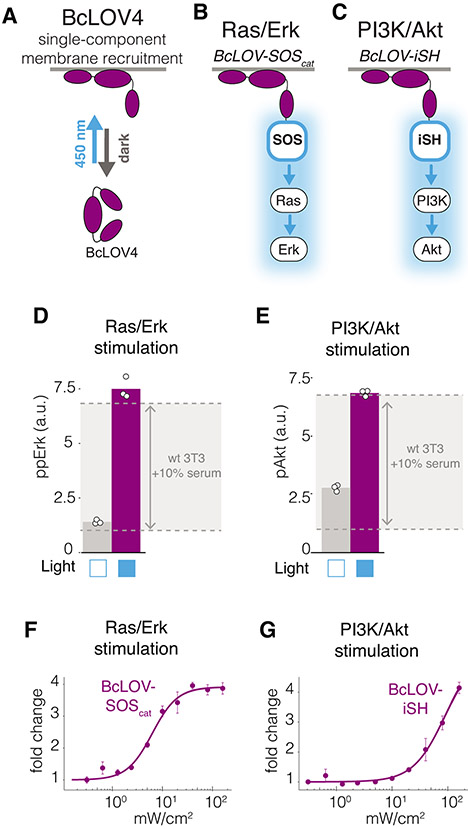
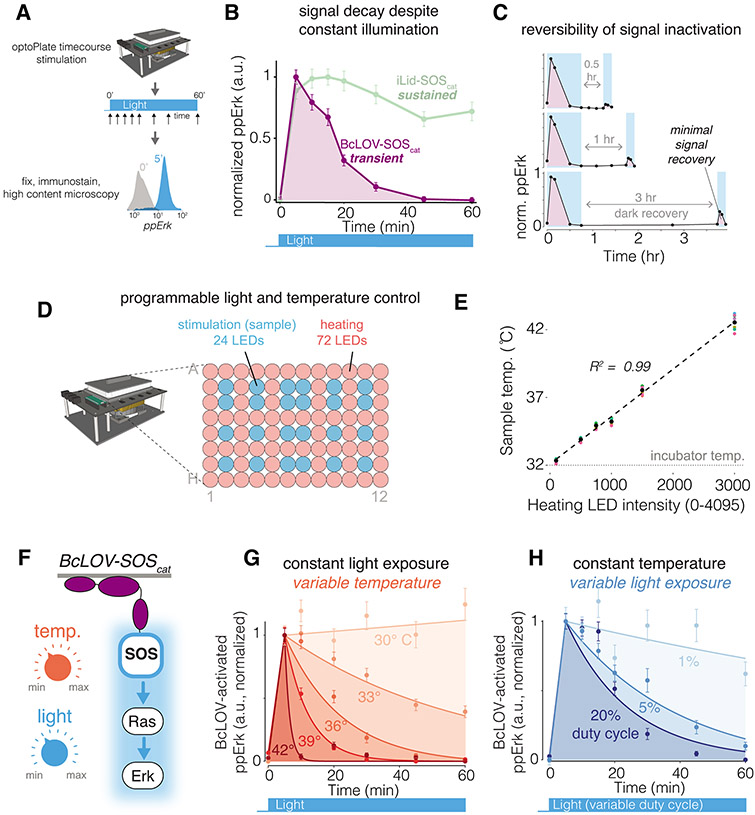
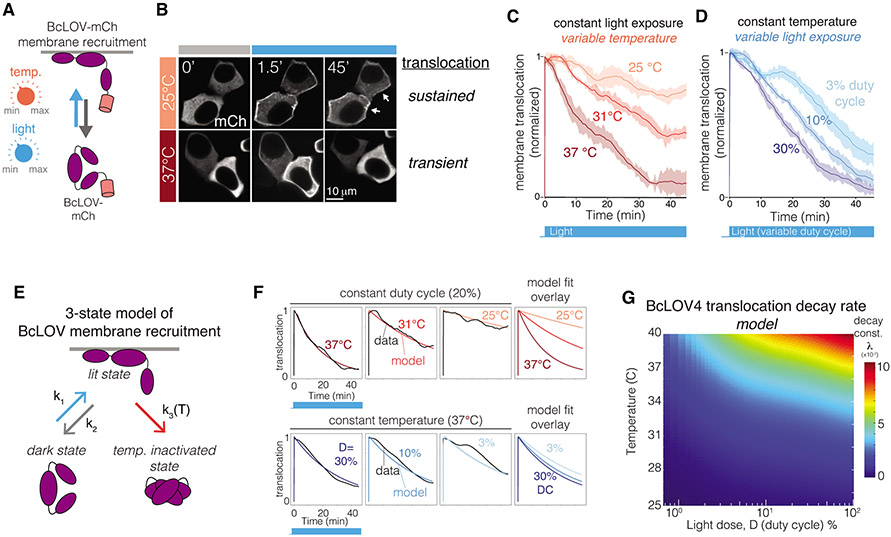
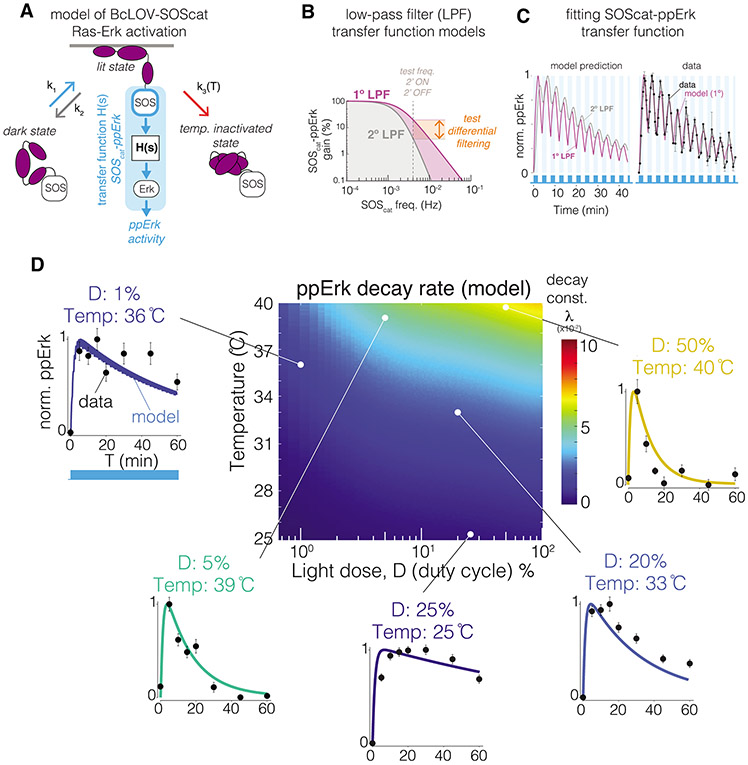
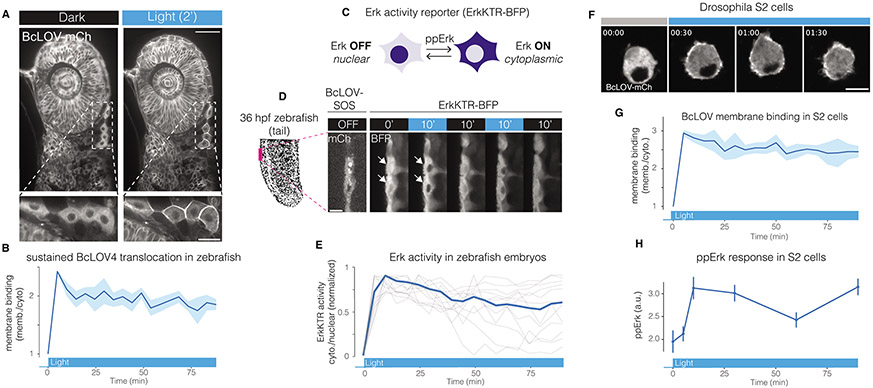
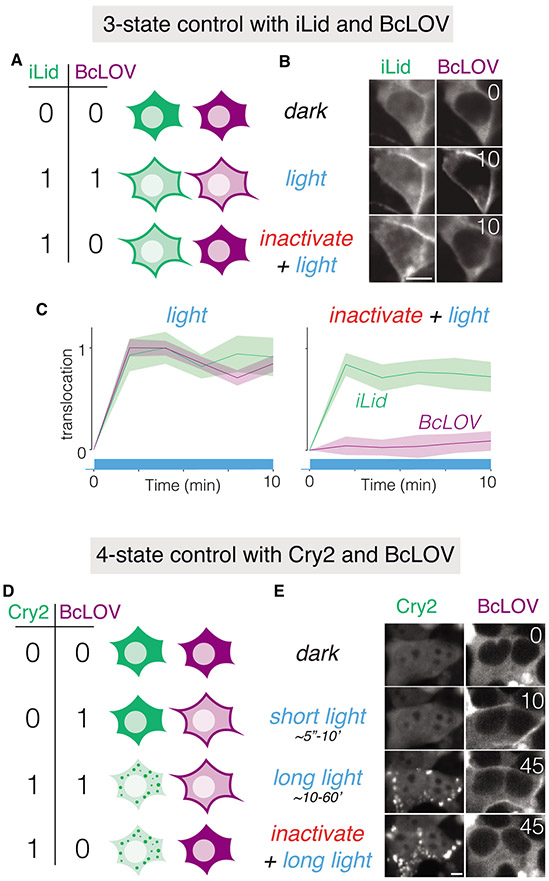
We describe single-component optogenetic probes whose activation dynamics depend on both light and temperature. We used the BcLOV4 photoreceptor to stimulate Ras and phosphatidyl inositol-3-kinase signaling in mammalian cells, allowing activation over a large dynamic range with low basal levels. Surprisingly, we found that BcLOV4 membrane translocation dynamics could be tuned by both light and temperature such that membrane localization spontaneously decayed at elevated temperatures despite constant illumination. Quantitative modeling predicted BcLOV4 activation dynamics across a range of light and temperature inputs and thus provides an experimental roadmap for BcLOV4-based probes. BcLOV4 drove strong and stable signal activation in both zebrafish and fly cells, and thermal inactivation provided a means to multiplex distinct blue-light sensitive tools in individual mammalian cells. BcLOV4 is thus a versatile photosensor with unique light and temperature sensitivity that enables straightforward generation of broadly applicable optogenetic tools.
© 2021. The Author(s), under exclusive licence to Springer Nature America, Inc.
Figures
Comment in
-
Optogenetics in the hot seat.Nat Chem Biol. 2022 Feb;18(2):118-119. doi: 10.1038/s41589-021-00928-x. Nat Chem Biol. 2022. PMID: 34937908 No abstract available.
References
Methods References
-
- Lamprecht MR, Sabatini DM & Carpenter AE CellProfiler™: free, versatile software for automated biological image analysis. Biotechniques 42, 71–75 (2007). - PubMed
-
- Wickham H, François R, Henry L & Müller K dplyr: a grammar of data manipulation. R package version 0.8. 0.1 Retrieved January 13, 2020 (2019).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
