

Structure of Hsp90-Hsp70-Hop-GR reveals the Hsp90 client-loading mechanism
- PMID: 34937942
- PMCID: PMC9179170
- DOI: 10.1038/s41586-021-04252-1
Structure of Hsp90-Hsp70-Hop-GR reveals the Hsp90 client-loading mechanism
Abstract
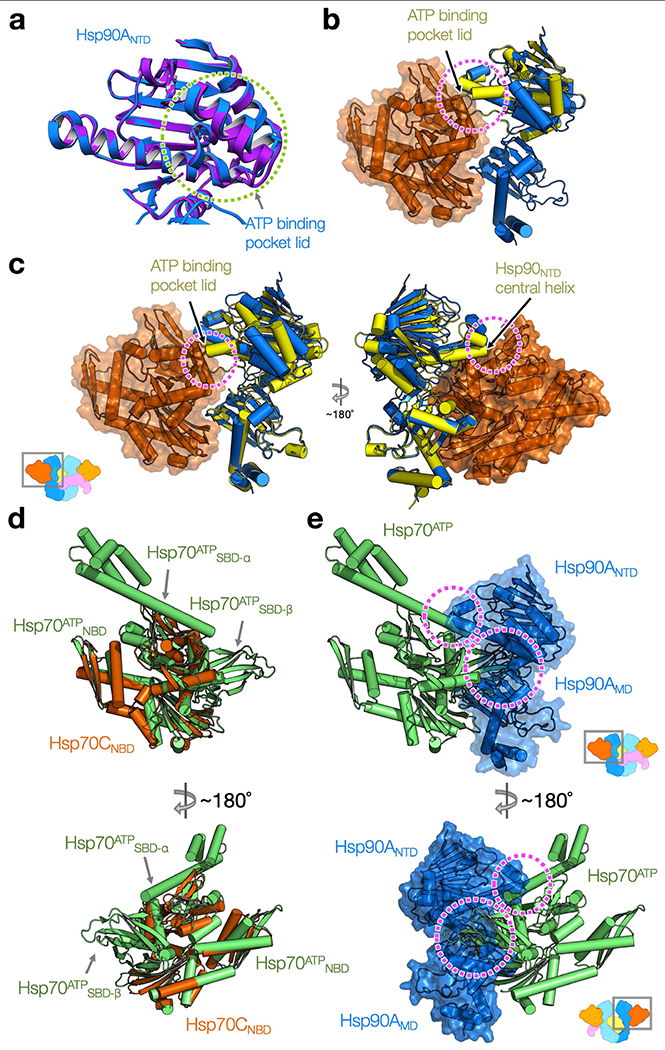
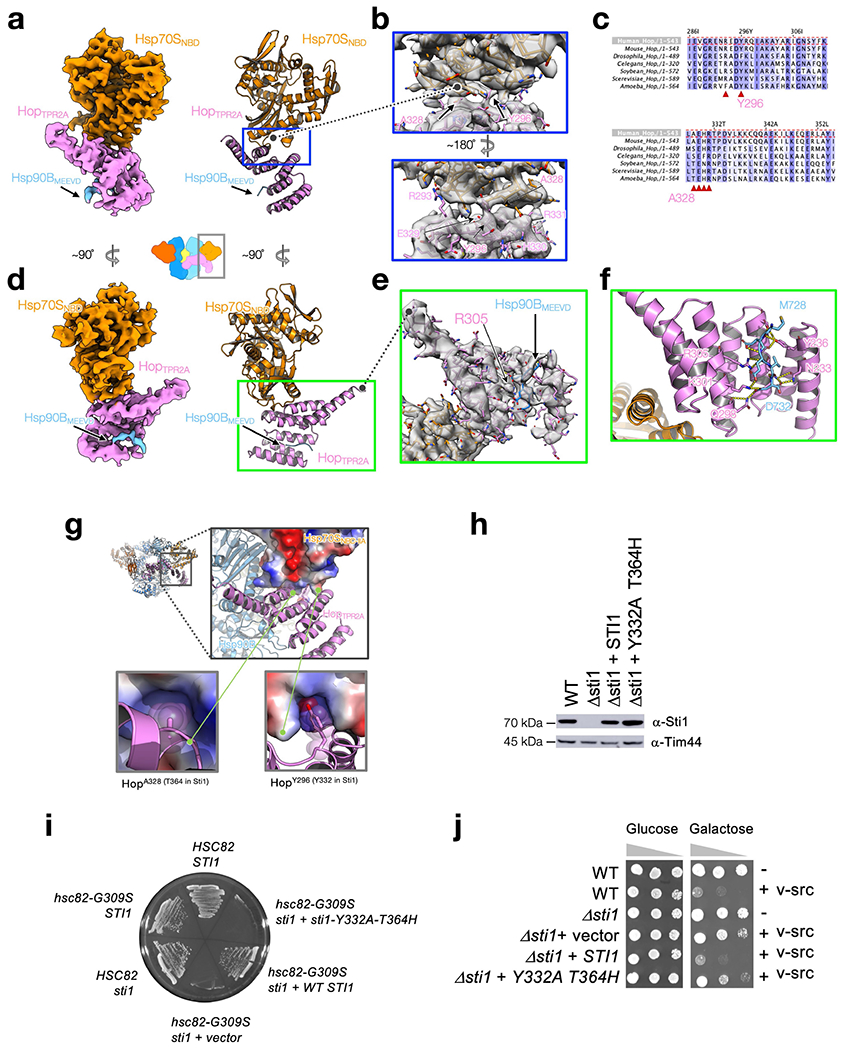
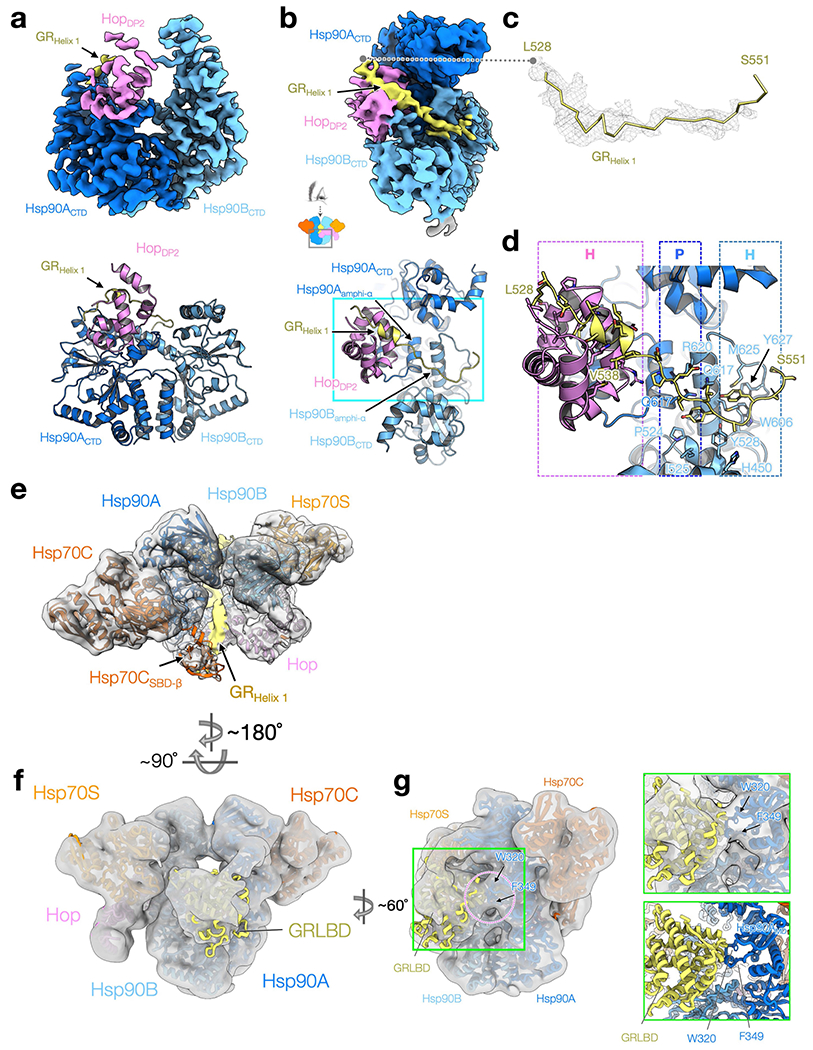
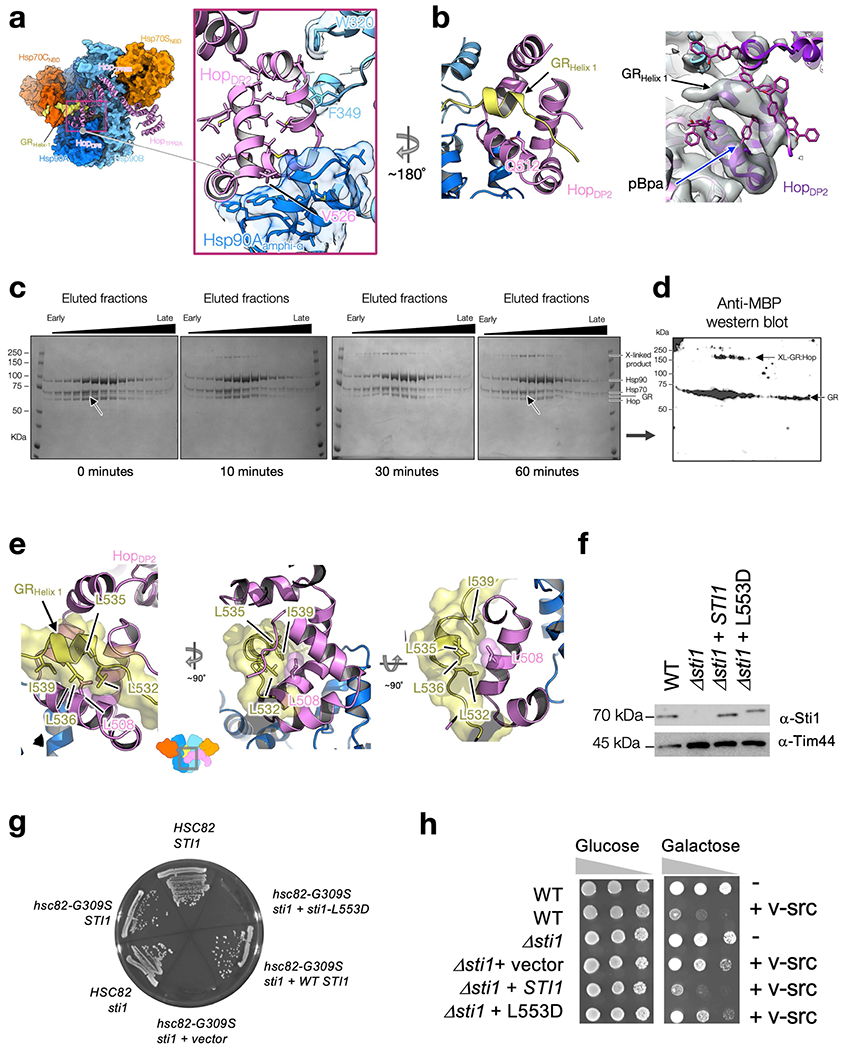
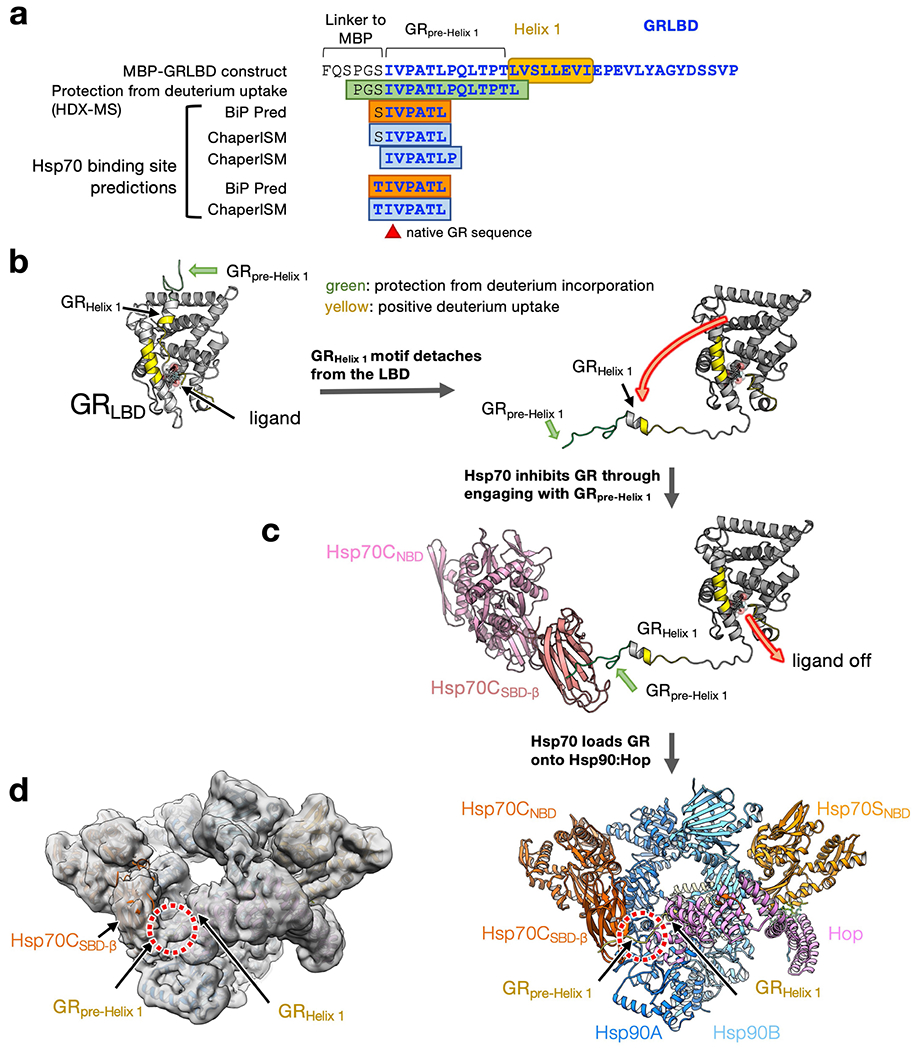
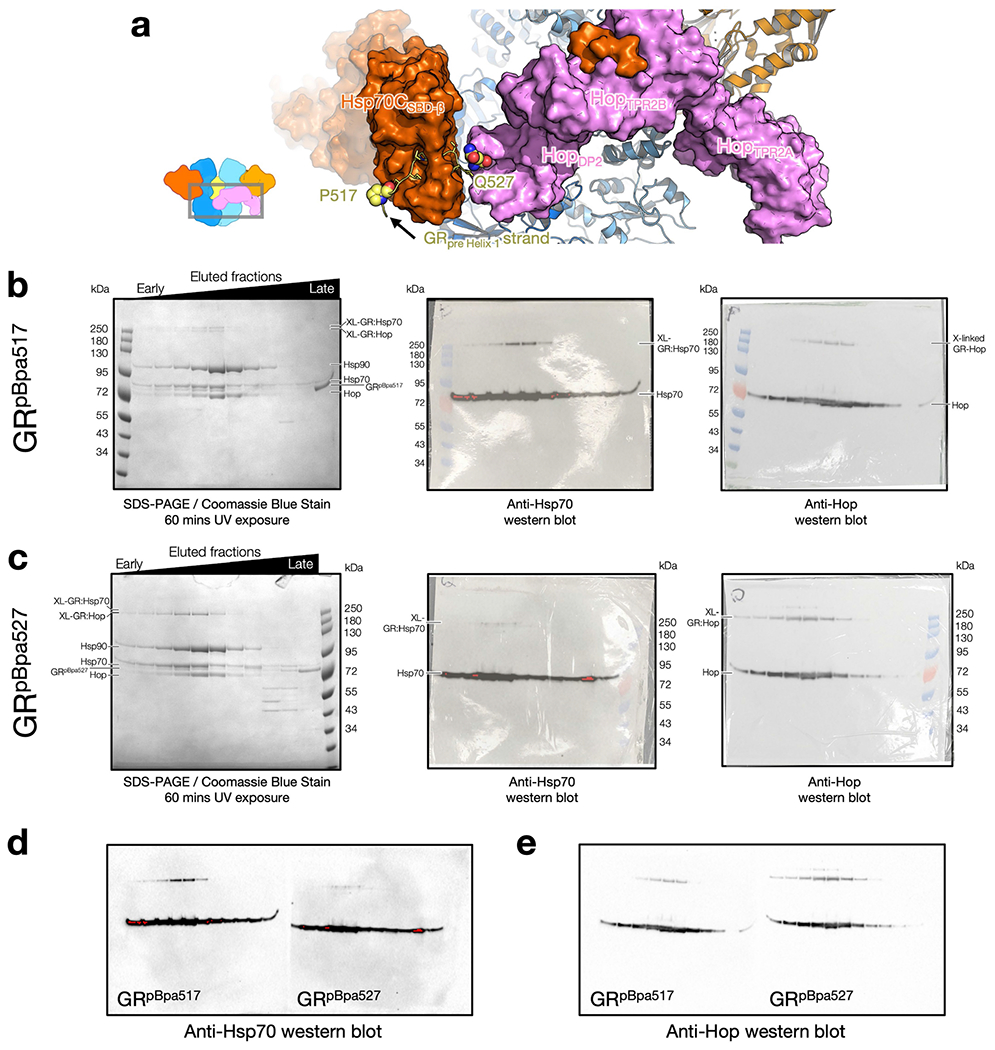
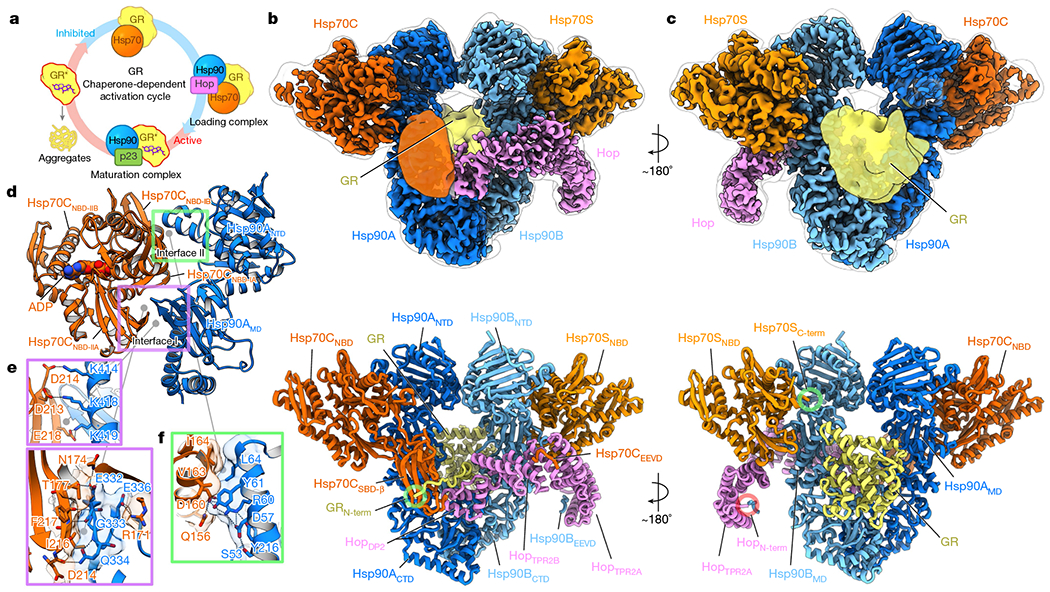
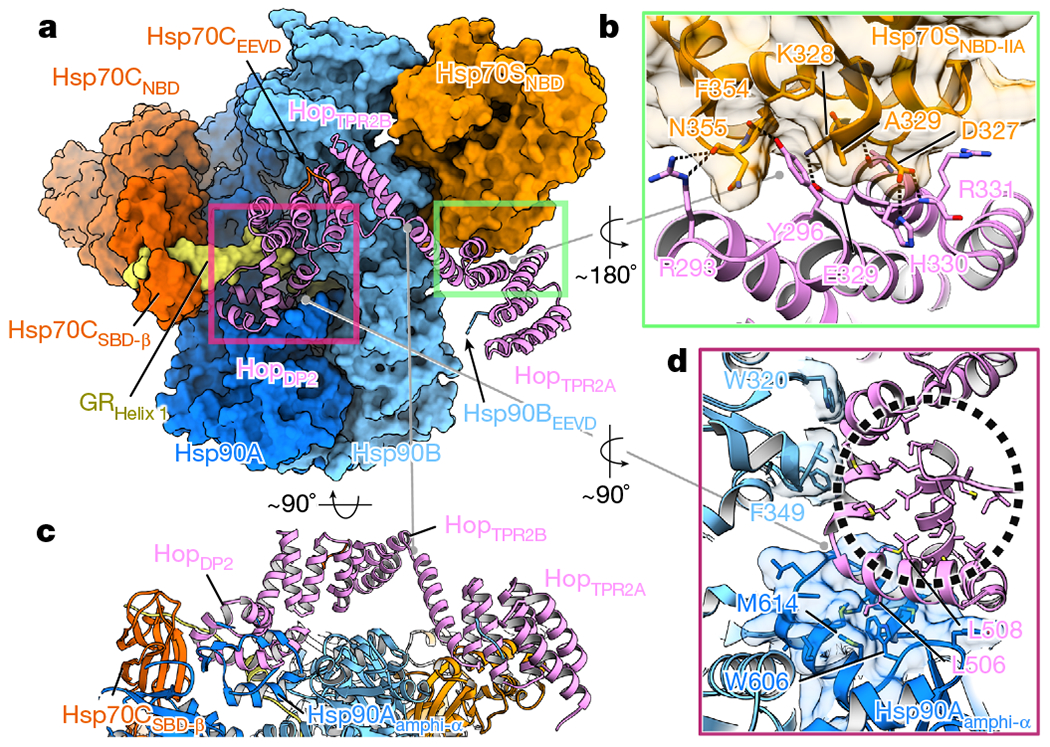
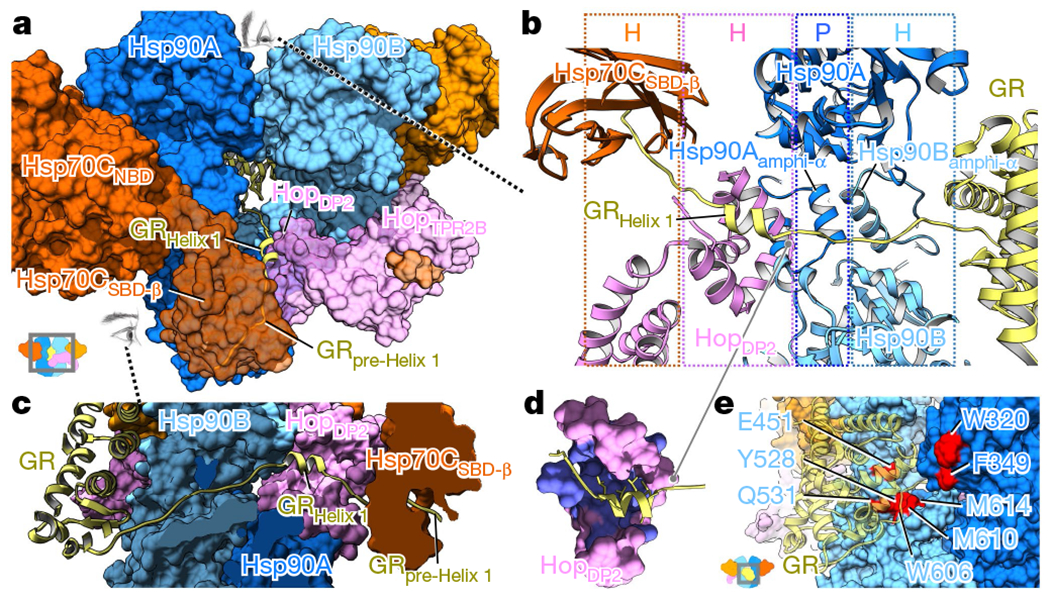
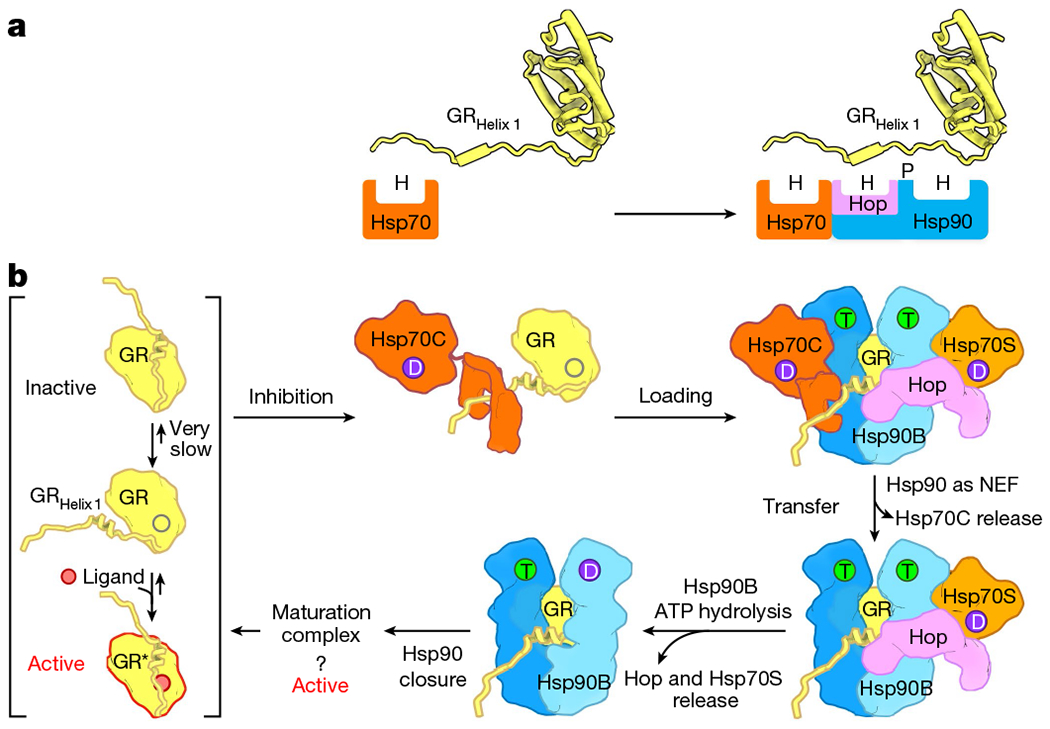
Maintaining a healthy proteome is fundamental for the survival of all organisms1. Integral to this are Hsp90 and Hsp70, molecular chaperones that together facilitate the folding, remodelling and maturation of the many 'client proteins' of Hsp902. The glucocorticoid receptor (GR) is a model client protein that is strictly dependent on Hsp90 and Hsp70 for activity3-7. Chaperoning GR involves a cycle of inactivation by Hsp70; formation of an inactive GR-Hsp90-Hsp70-Hop 'loading' complex; conversion to an active GR-Hsp90-p23 'maturation' complex; and subsequent GR release8. However, to our knowledge, a molecular understanding of this intricate chaperone cycle is lacking for any client protein. Here we report the cryo-electron microscopy structure of the GR-loading complex, in which Hsp70 loads GR onto Hsp90, uncovering the molecular basis of direct coordination by Hsp90 and Hsp70. The structure reveals two Hsp70 proteins, one of which delivers GR and the other scaffolds the Hop cochaperone. Hop interacts with all components of the complex, including GR, and poises Hsp90 for subsequent ATP hydrolysis. GR is partially unfolded and recognized through an extended binding pocket composed of Hsp90, Hsp70 and Hop, revealing the mechanism of GR loading and inactivation. Together with the GR-maturation complex structure9, we present a complete molecular mechanism of chaperone-dependent client remodelling, and establish general principles of client recognition, inhibition, transfer and activation.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures
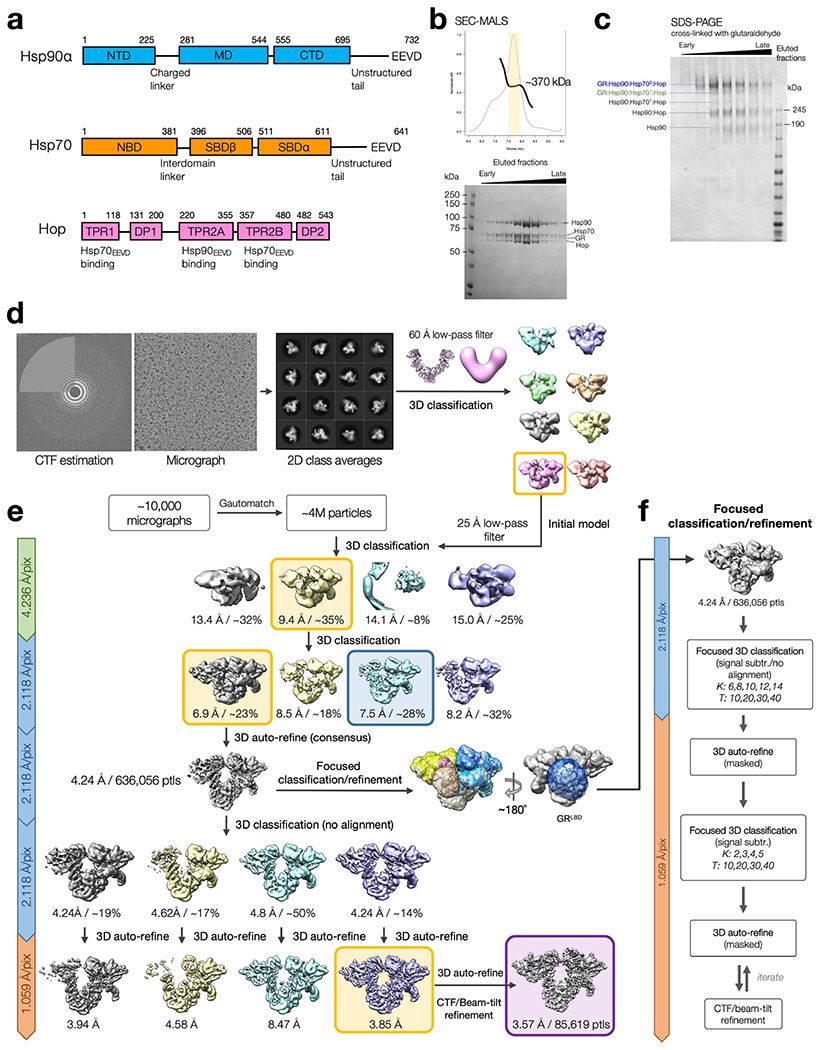
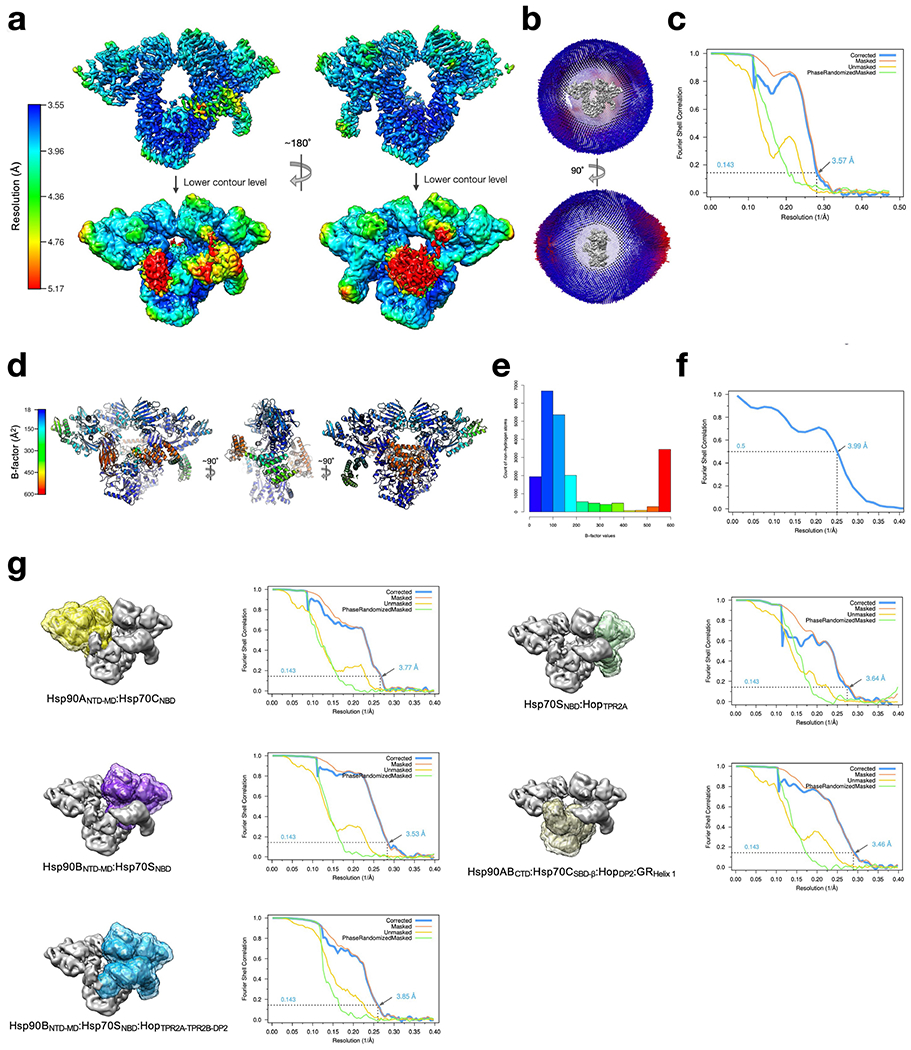
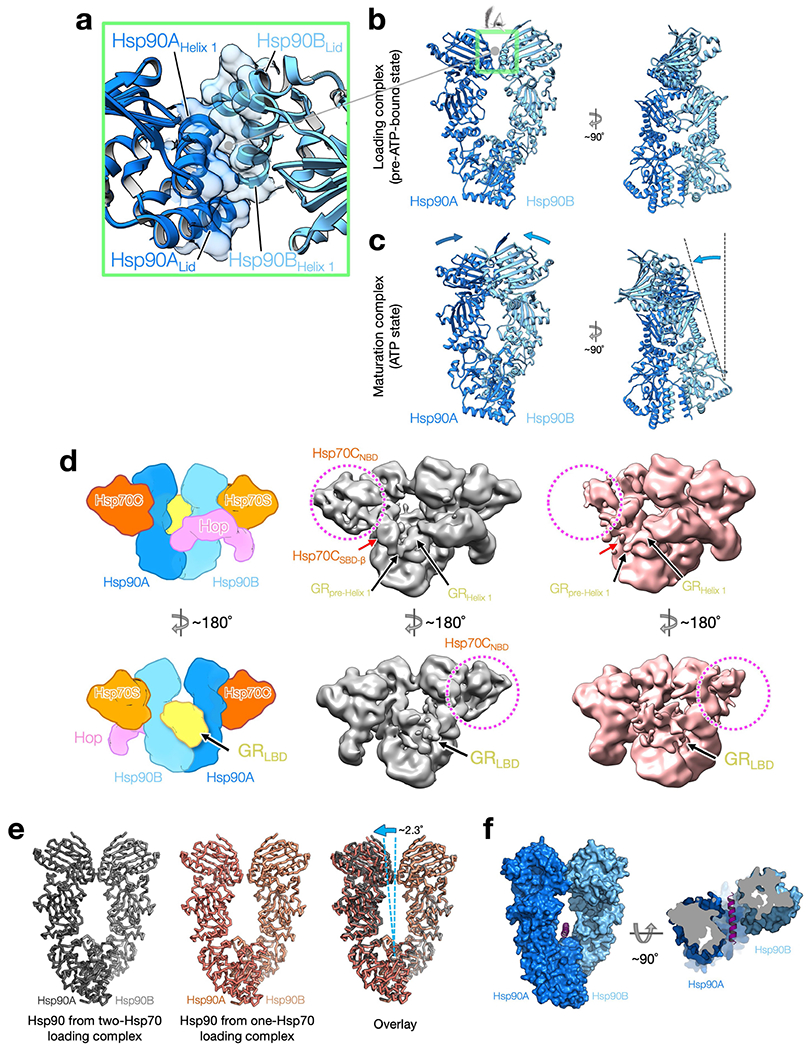
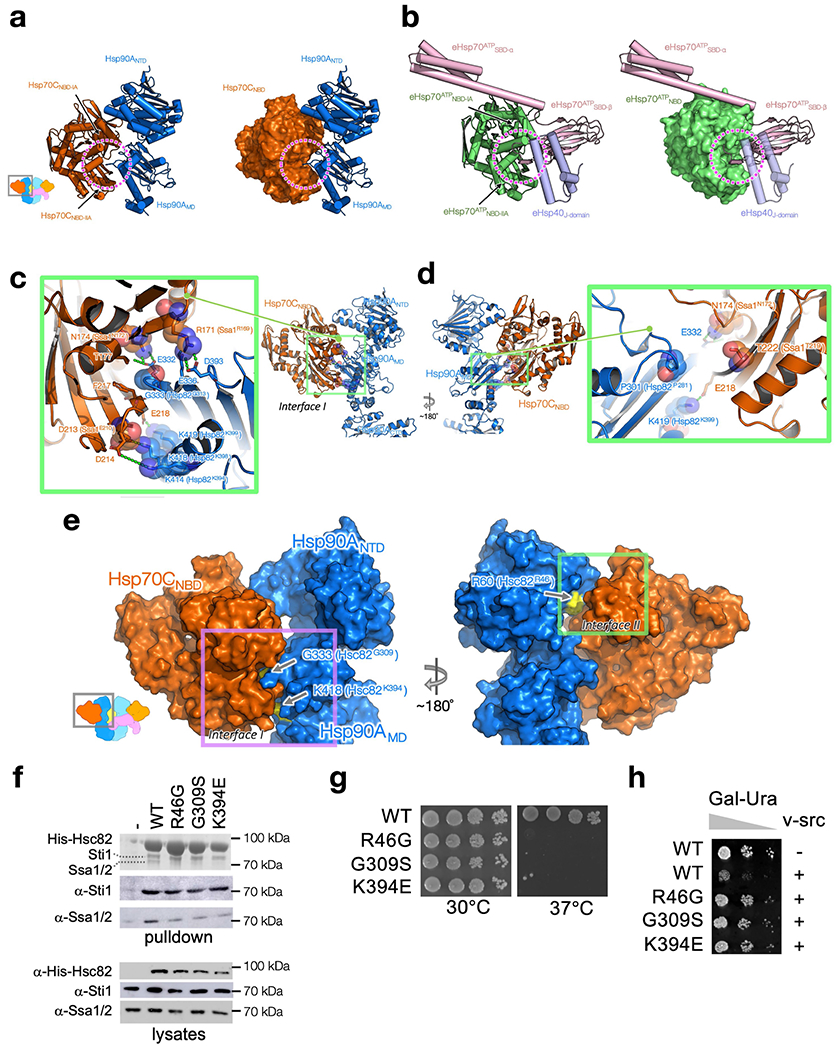
References
-
- Kim YE, Hipp MS, Bracher A, Hayer-Hartl M & Hartl FU Molecular chaperone functions in protein folding and proteostasis. Annu. Rev. Biochem 82, 323–355 (2013). - PubMed
-
- Lorenz OR et al. Modulation of the Hsp90 chaperone cycle by a stringent client protein. Mol. Cell 53, 941–953 (2014). - PubMed
-
- Picard D et al. Reduced levels of hsp90 compromise steroid receptor action in vivo. Nature 348, 166–168 (1990). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
