

Valine tRNA levels and availability regulate complex I assembly in leukaemia
- PMID: 34937946
- PMCID: PMC9116157
- DOI: 10.1038/s41586-021-04244-1
Valine tRNA levels and availability regulate complex I assembly in leukaemia
Abstract
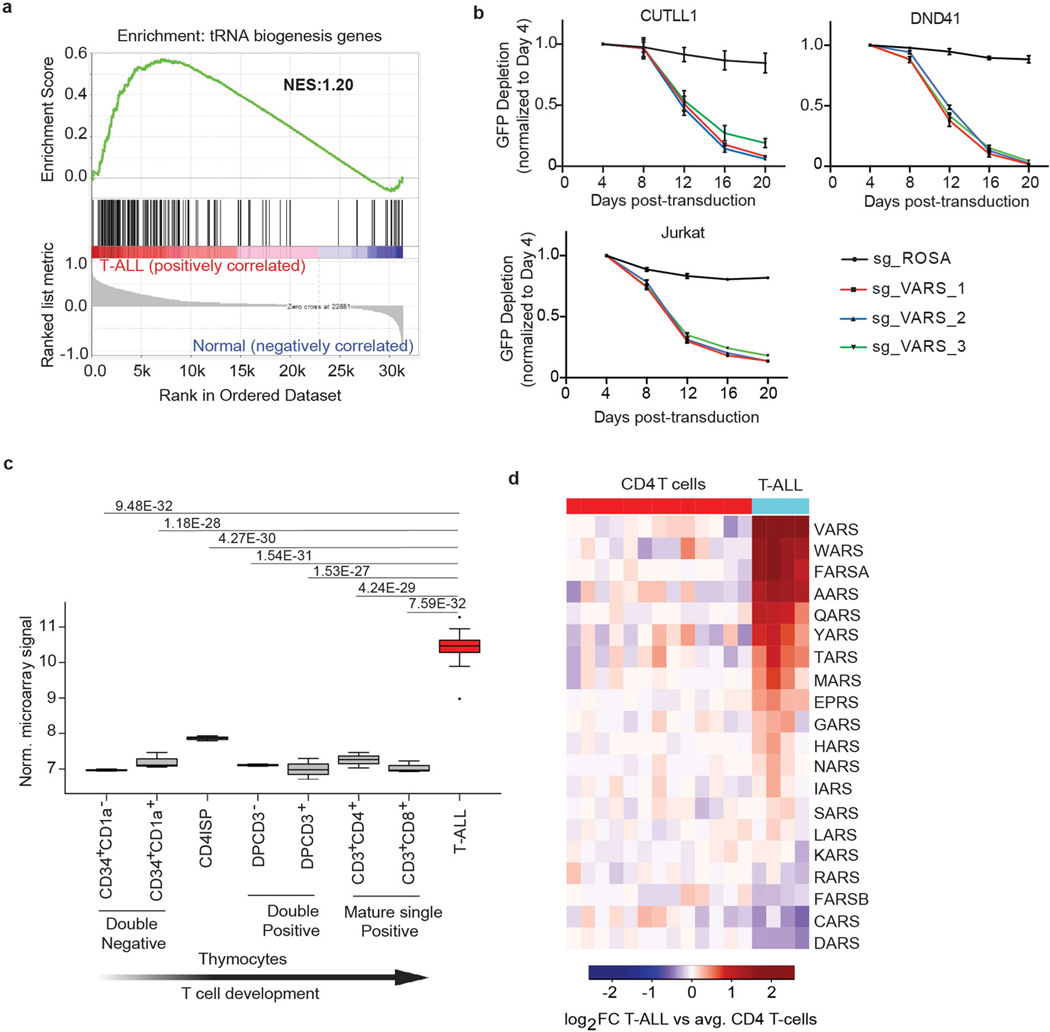
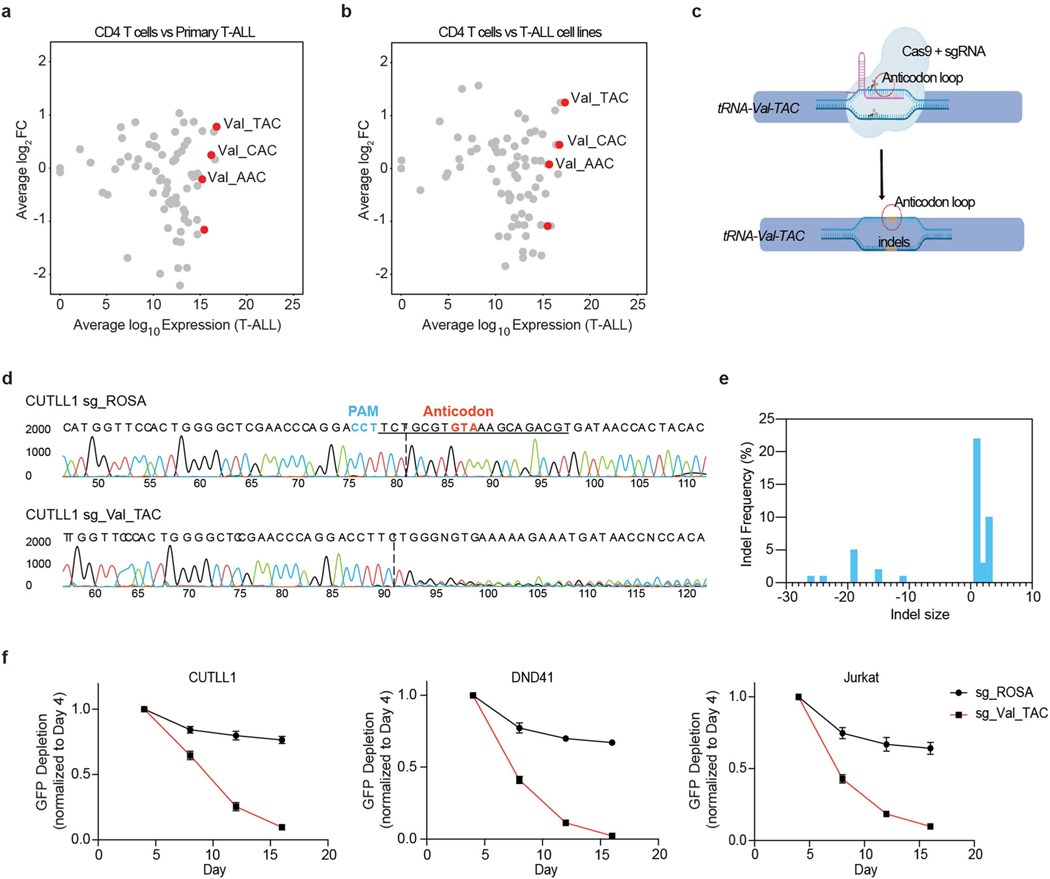
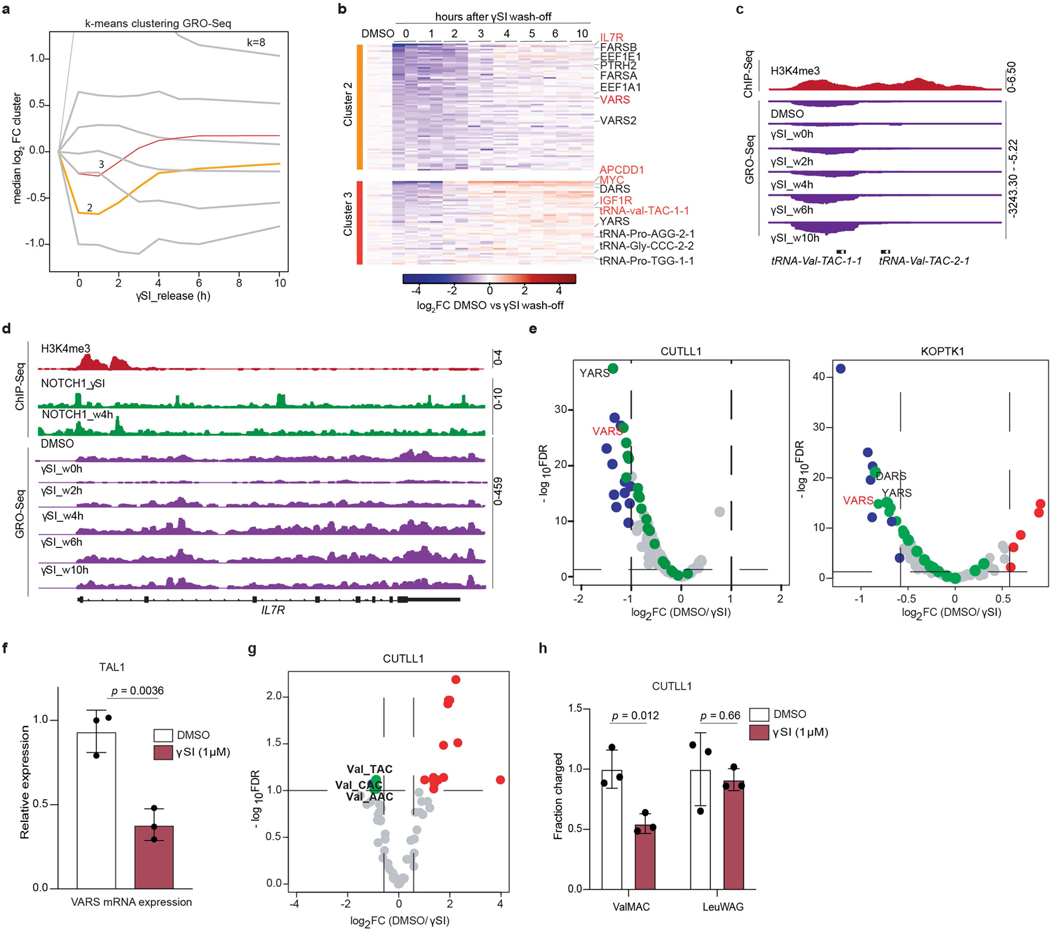
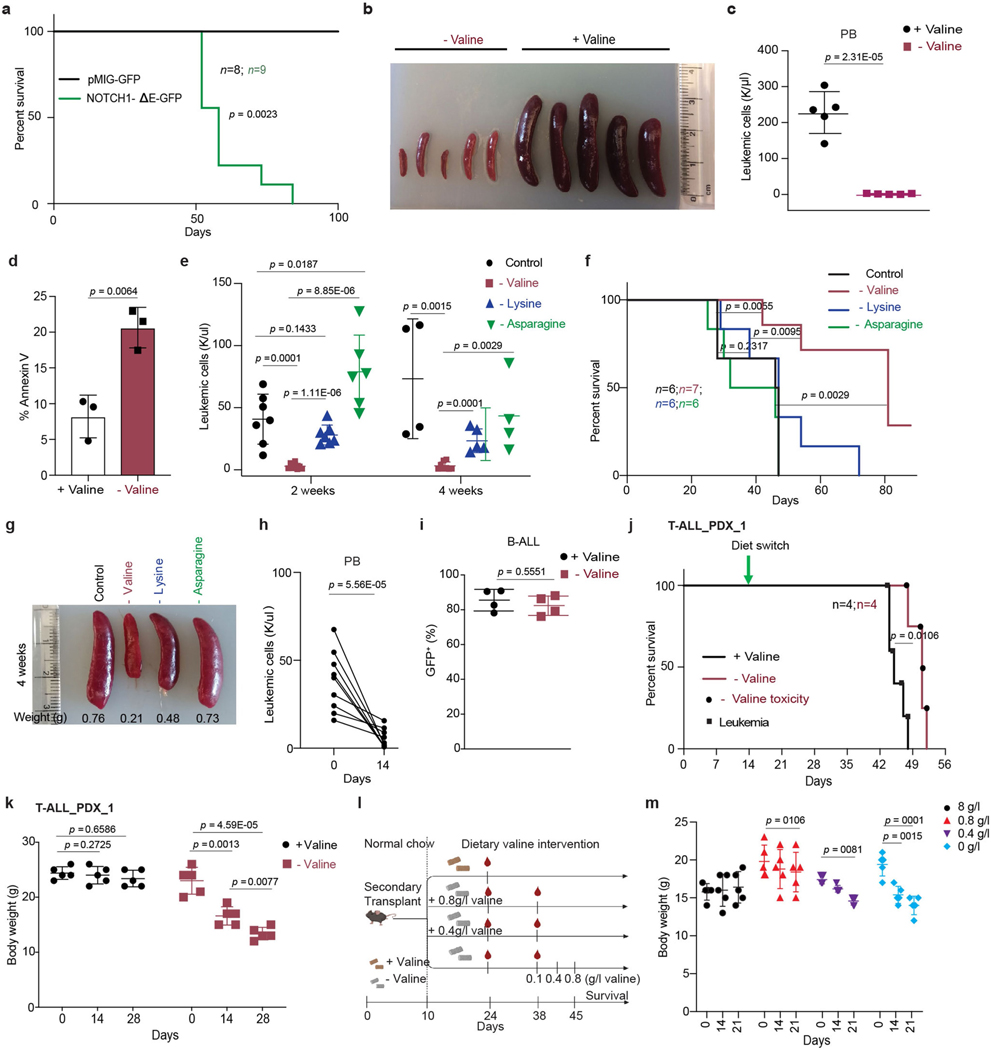
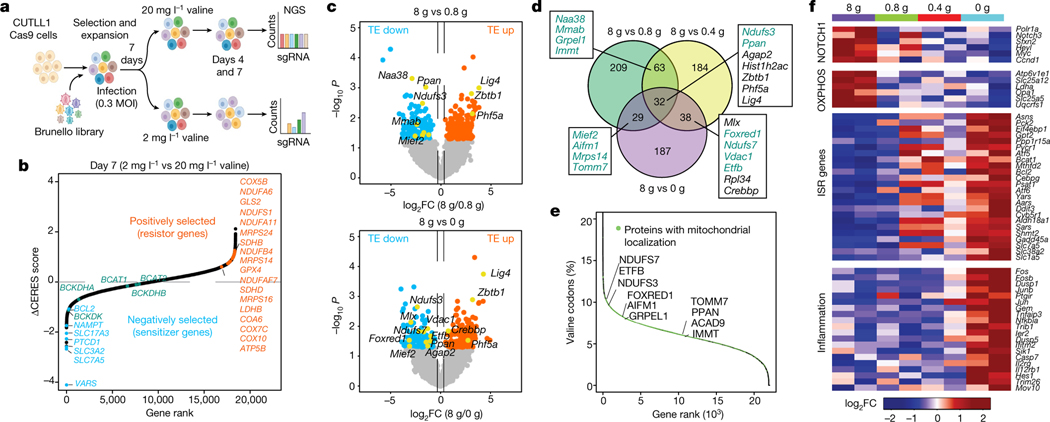
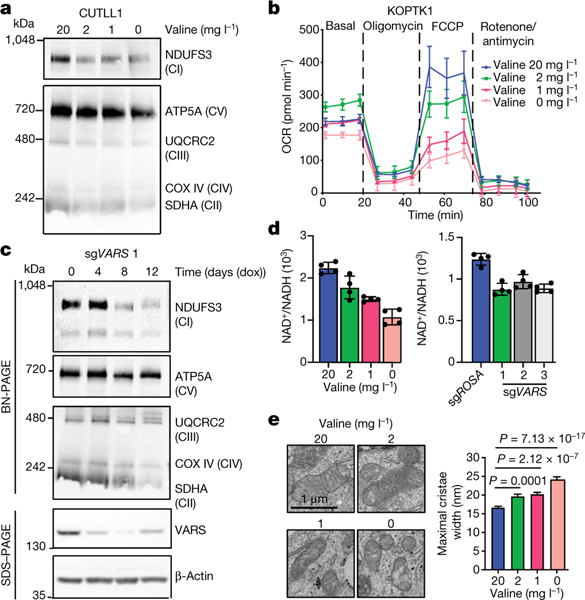
Although deregulation of transfer RNA (tRNA) biogenesis promotes the translation of pro-tumorigenic mRNAs in cancers1,2, the mechanisms and consequences of tRNA deregulation in tumorigenesis are poorly understood. Here we use a CRISPR-Cas9 screen to focus on genes that have been implicated in tRNA biogenesis, and identify a mechanism by which altered valine tRNA biogenesis enhances mitochondrial bioenergetics in T cell acute lymphoblastic leukaemia (T-ALL). Expression of valine aminoacyl tRNA synthetase is transcriptionally upregulated by NOTCH1, a key oncogene in T-ALL, underlining a role for oncogenic transcriptional programs in coordinating tRNA supply and demand. Limiting valine bioavailability through restriction of dietary valine intake disrupted this balance in mice, resulting in decreased leukaemic burden and increased survival in vivo. Mechanistically, valine restriction reduced translation rates of mRNAs that encode subunits of mitochondrial complex I, leading to defective assembly of complex I and impaired oxidative phosphorylation. Finally, a genome-wide CRISPR-Cas9 loss-of-function screen in differential valine conditions identified several genes, including SLC7A5 and BCL2, whose genetic ablation or pharmacological inhibition synergized with valine restriction to reduce T-ALL growth. Our findings identify tRNA deregulation as a critical adaptation in the pathogenesis of T-ALL and provide a molecular basis for the use of dietary approaches to target tRNA biogenesis in blood malignancies.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
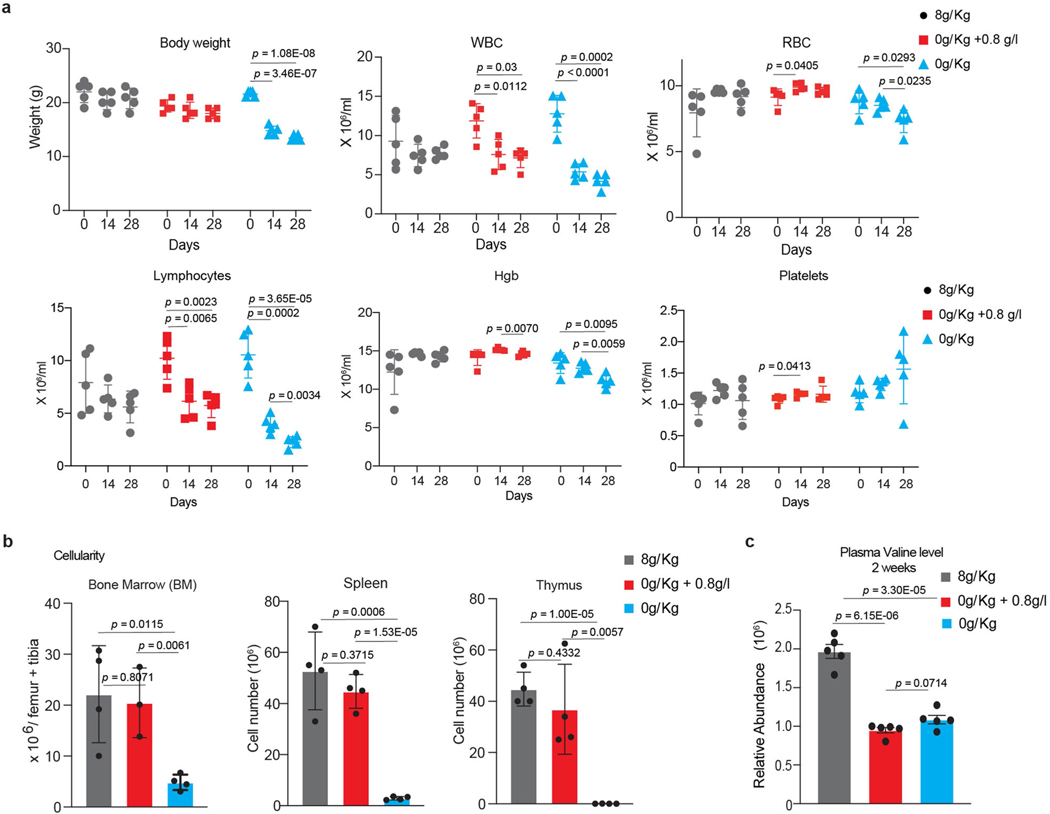
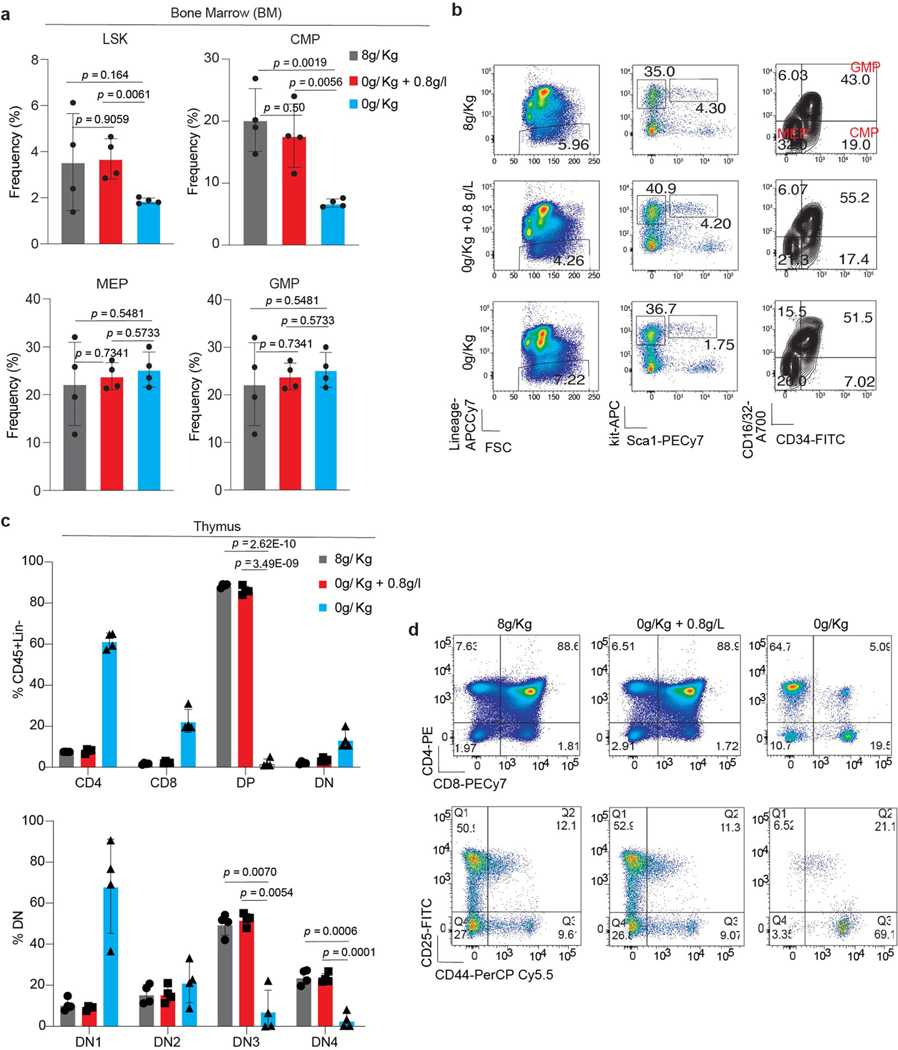
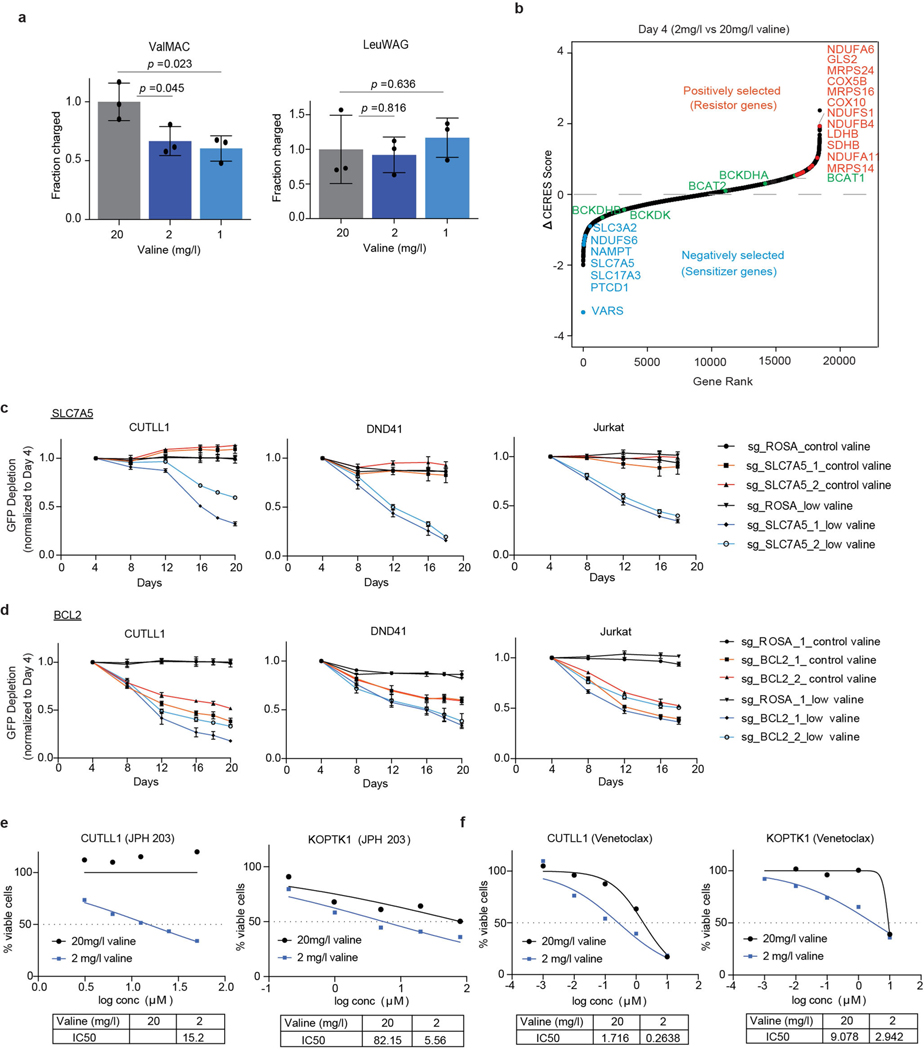
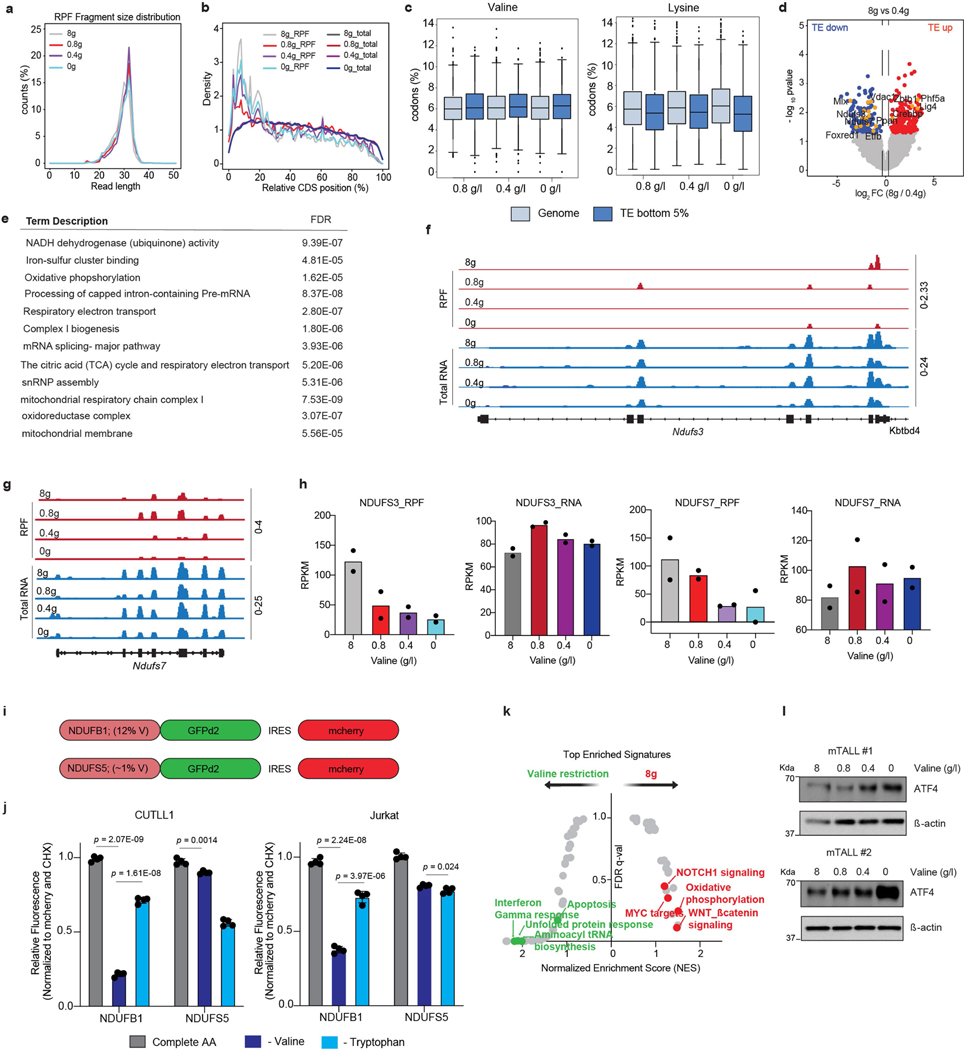
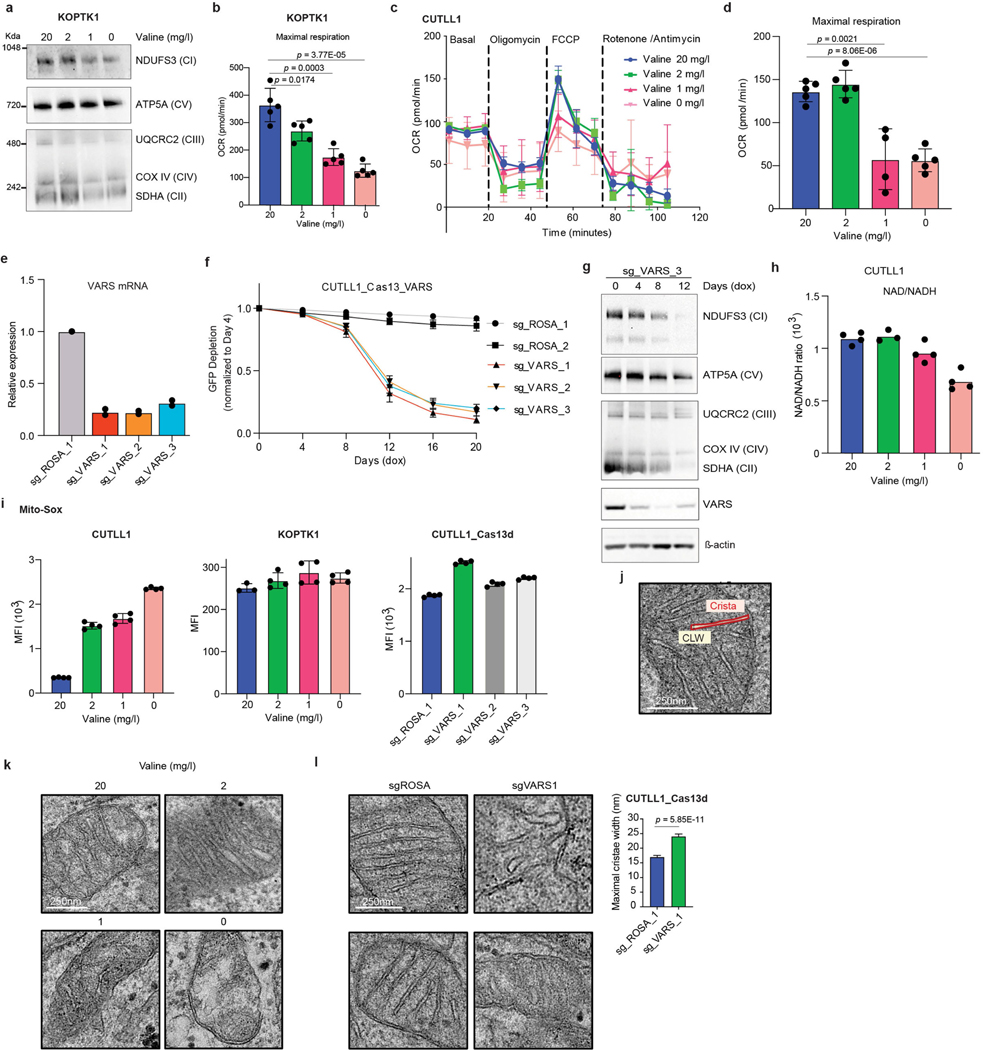
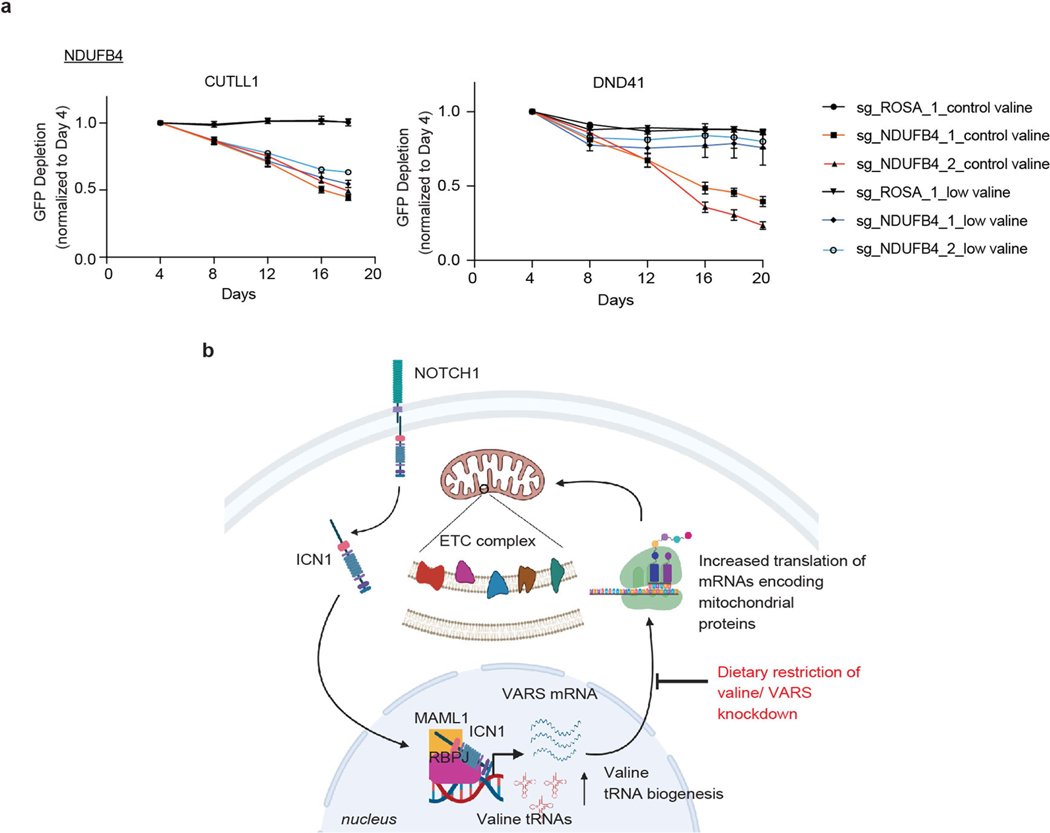
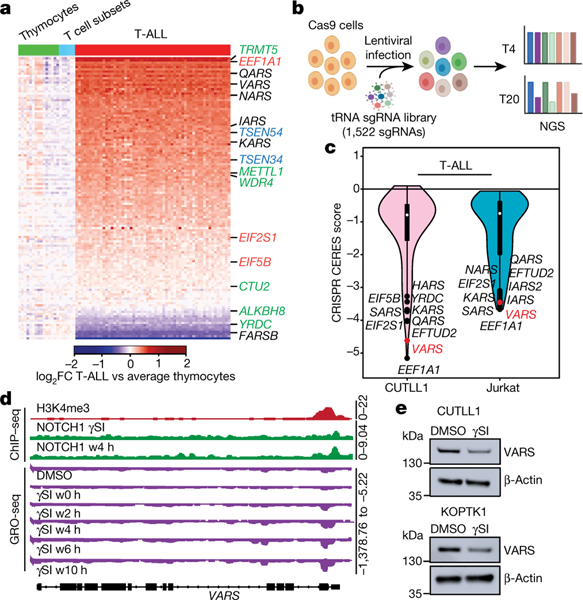
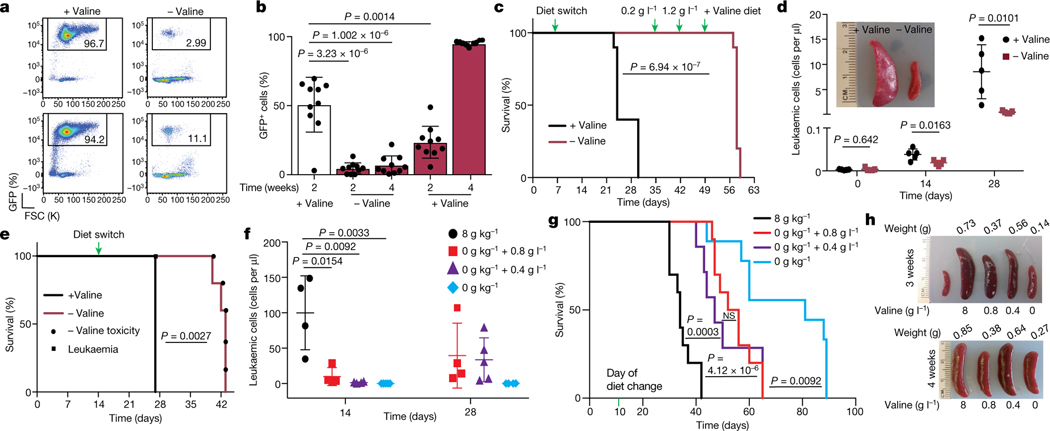
Comment in
-
tRNA supply and demand.Nat Rev Cancer. 2022 Mar;22(3):129. doi: 10.1038/s41568-022-00448-3. Nat Rev Cancer. 2022. PMID: 35091697 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA173636/CA/NCI NIH HHS/United States
- P01 CA229086/CA/NCI NIH HHS/United States
- P30 CA016087/CA/NCI NIH HHS/United States
- R01 CA252239/CA/NCI NIH HHS/United States
- R01 CA228135/CA/NCI NIH HHS/United States
- R01 CA227649/CA/NCI NIH HHS/United States
- K22 CA201088/CA/NCI NIH HHS/United States
- R01 CA266212/CA/NCI NIH HHS/United States
- R01 CA216421/CA/NCI NIH HHS/United States
- R01 HL159175/HL/NHLBI NIH HHS/United States
- R01 CA242020/CA/NCI NIH HHS/United States
- K99 CA252602/CA/NCI NIH HHS/United States
- R37 CA222504/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
