

Cell-by-cell dissection of phloem development links a maturation gradient to cell specialization
- PMID: 34941412
- PMCID: PMC8730638
- DOI: 10.1126/science.aba5531
Cell-by-cell dissection of phloem development links a maturation gradient to cell specialization
Abstract
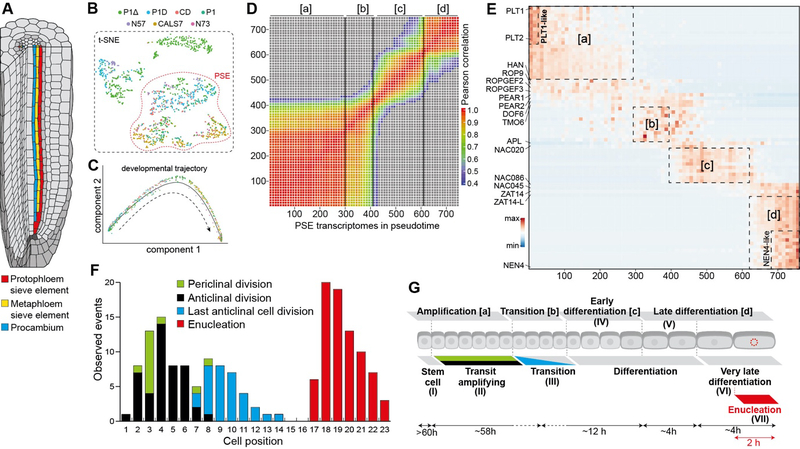
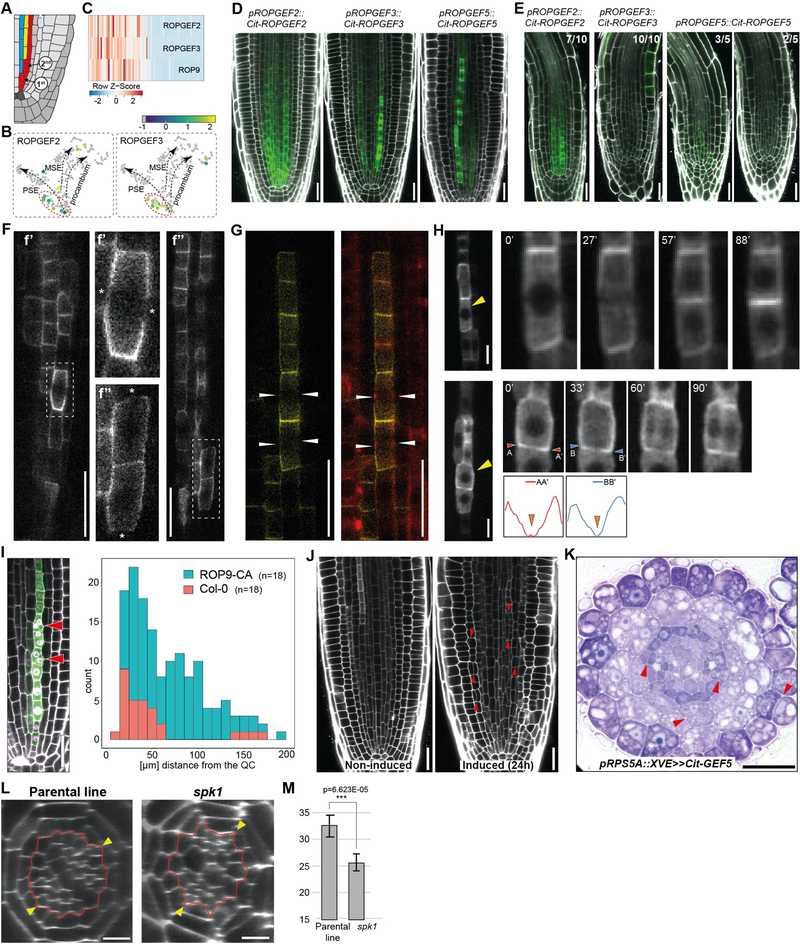
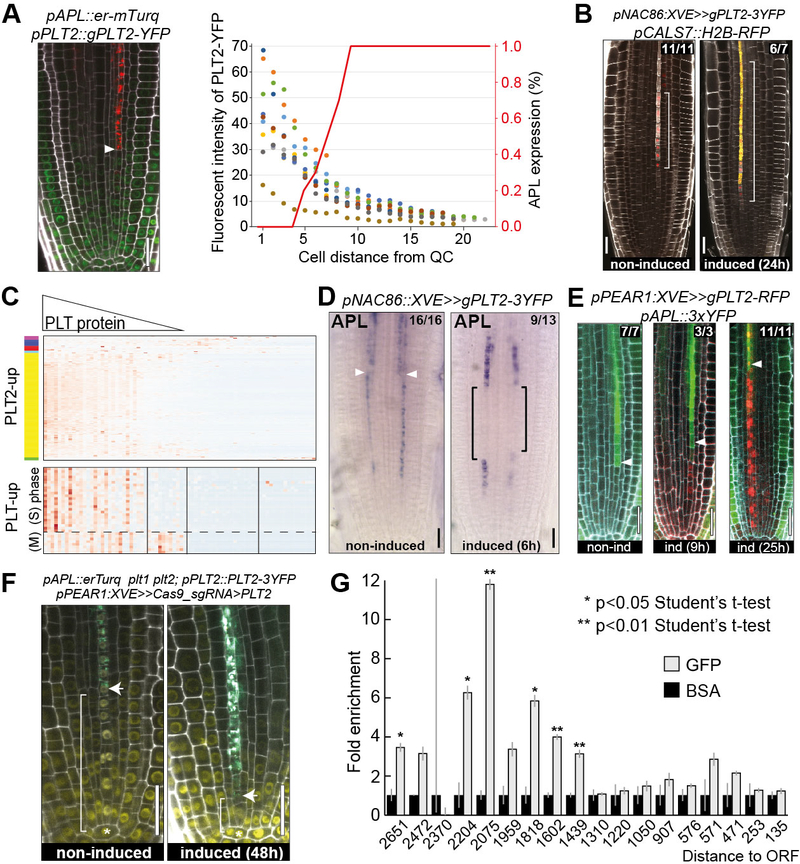
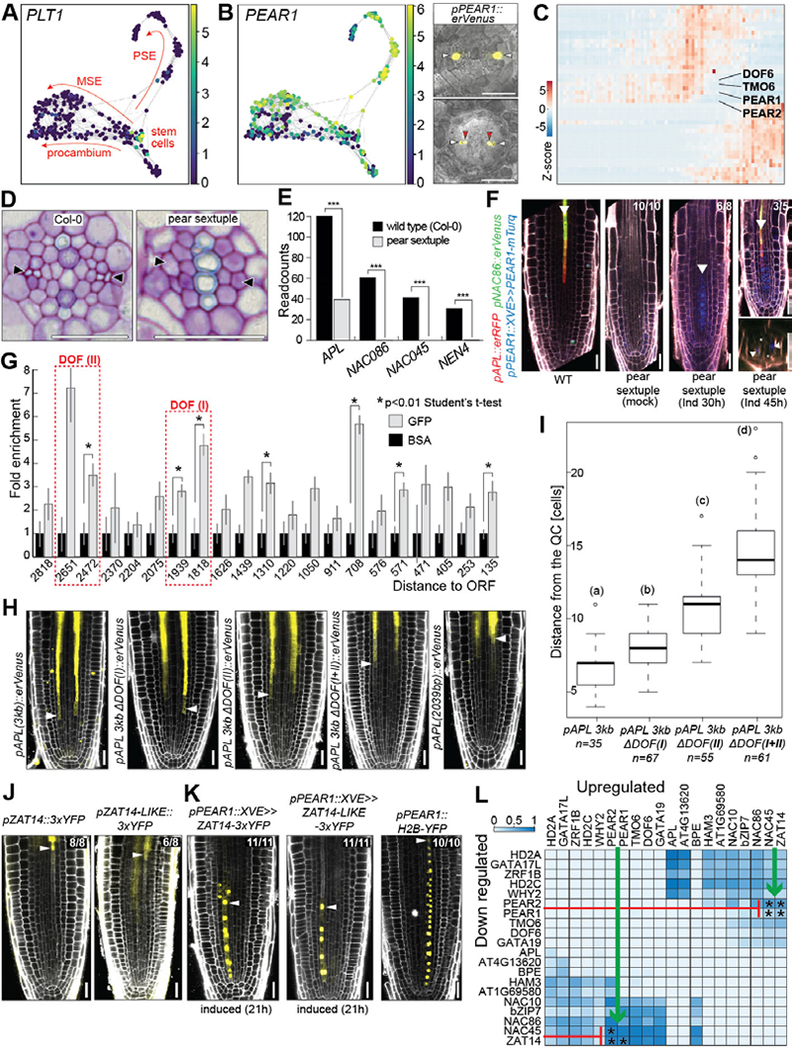
In the plant meristem, tissue-wide maturation gradients are coordinated with specialized cell networks to establish various developmental phases required for indeterminate growth. Here, we used single-cell transcriptomics to reconstruct the protophloem developmental trajectory from the birth of cell progenitors to terminal differentiation in the Arabidopsis thaliana root. PHLOEM EARLY DNA-BINDING-WITH-ONE-FINGER (PEAR) transcription factors mediate lineage bifurcation by activating guanosine triphosphatase signaling and prime a transcriptional differentiation program. This program is initially repressed by a meristem-wide gradient of PLETHORA transcription factors. Only the dissipation of PLETHORA gradient permits activation of the differentiation program that involves mutual inhibition of early versus late meristem regulators. Thus, for phloem development, broad maturation gradients interface with cell-type-specific transcriptional regulators to stage cellular differentiation.
Conflict of interest statement
Figures
References
-
- Dolan L, Janmaat K, Willemsen V, Linstead P, Poethig S, Roberts K, Scheres B, Cellular organisation of the Arabidopsis thaliana root. Development. 119, 71–84 (1993). - PubMed
-
- Furuta KM, Yadav SR, Lehesranta S, Belevich I, Miyashima S, Heo J, Vatén A, Lindgren O, Rybel BD, Isterdael GV, Somervuo P, Lichtenberger R, Rocha R, Thitamadee S, Tähtiharju S, Auvinen P, Beeckman T, Jokitalo E, Helariutta Y, Arabidopsis NAC45/86 direct sieve element morphogenesis culminating in enucleation. Science. 345, 933–937 (2014). - PubMed
-
- Marhava P, Bassukas AEL, Zourelidou M, Kolb M, Moret B, Fastner A, Schulze WX, Cattaneo P, Hammes UZ, Schwechheimer C, Hardtke CS, A molecular rheostat adjusts auxin flux to promote root protophloem differentiation. Nature. 558, 297–300 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
