

Broadly neutralizing antibodies target a haemagglutinin anchor epitope
- PMID: 34942633
- PMCID: PMC8828479
- DOI: 10.1038/s41586-021-04356-8
Broadly neutralizing antibodies target a haemagglutinin anchor epitope
Abstract
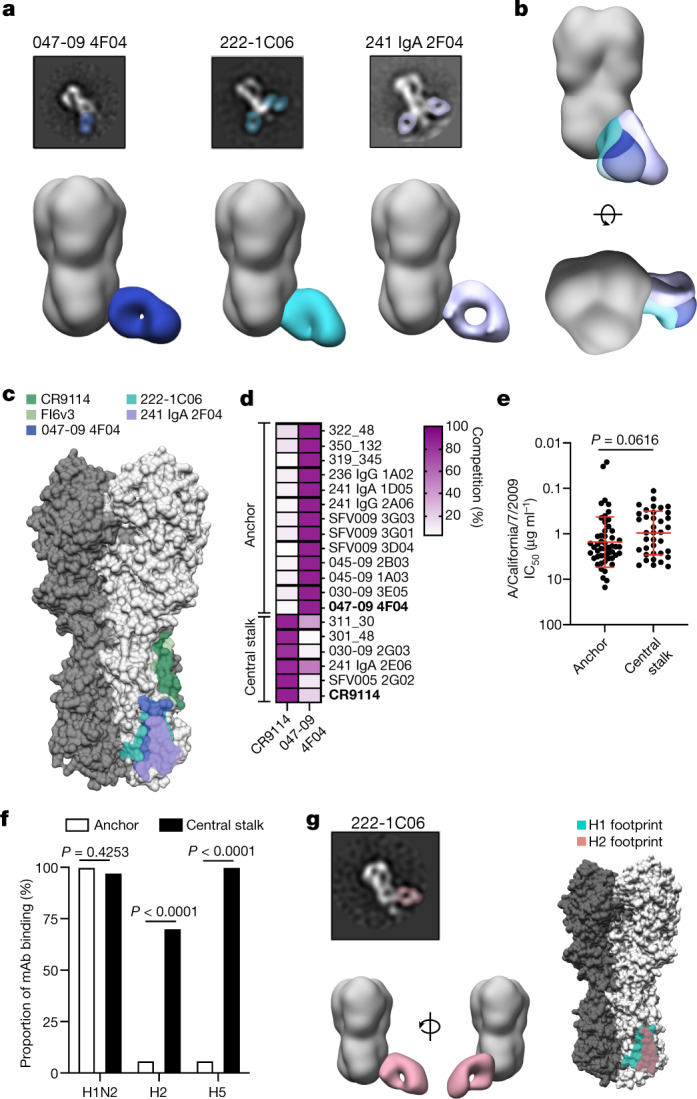
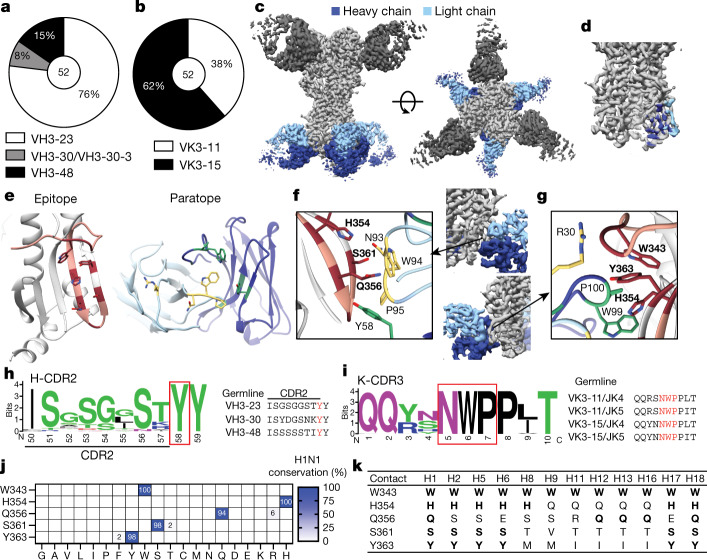
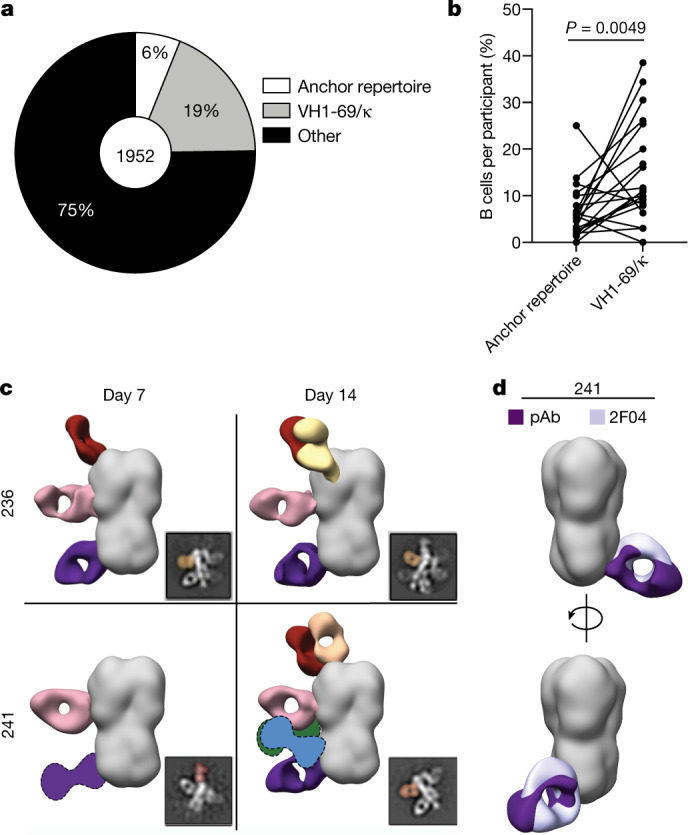
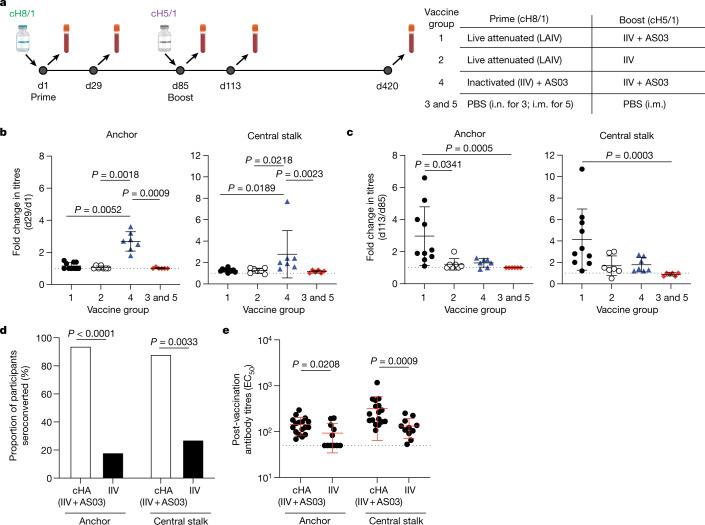
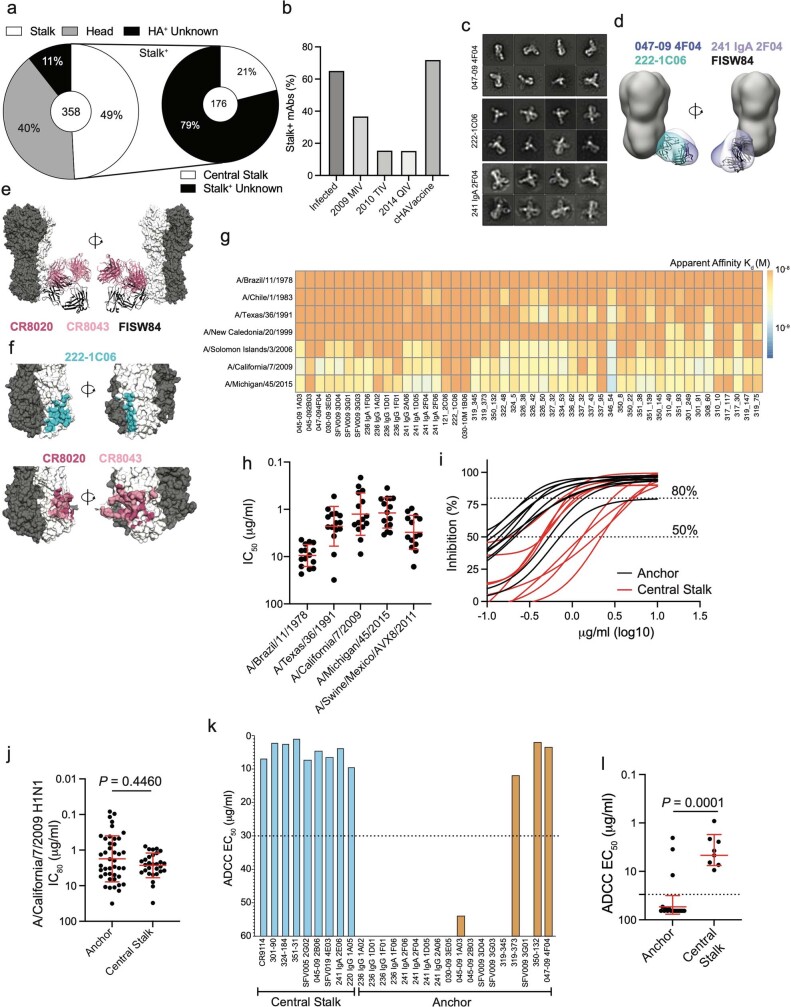
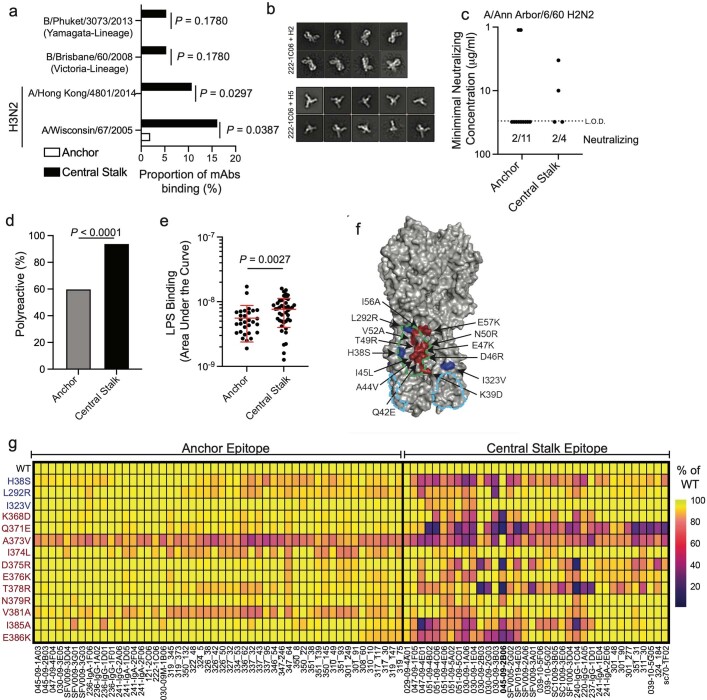
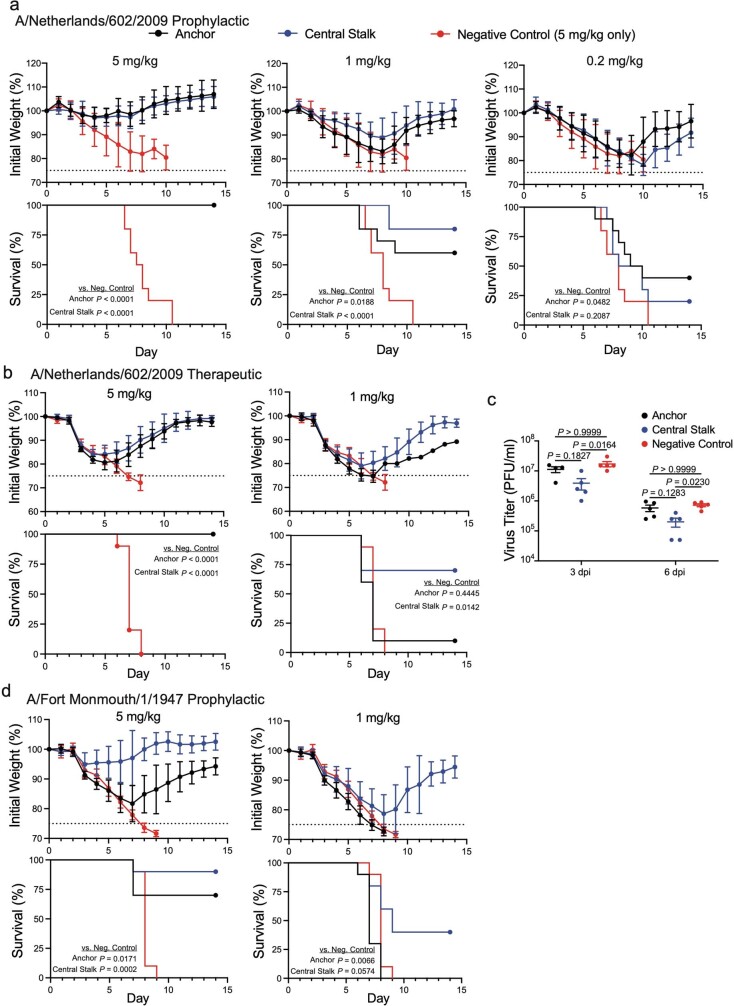
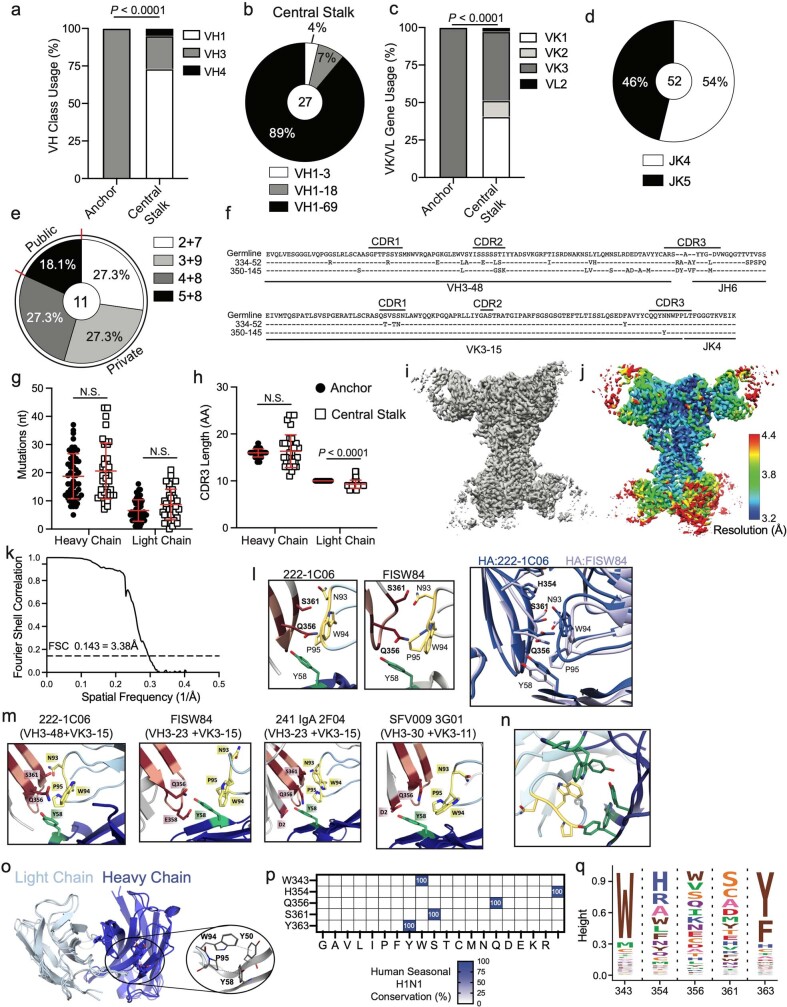
Broadly neutralizing antibodies that target epitopes of haemagglutinin on the influenza virus have the potential to provide near universal protection against influenza virus infection1. However, viral mutants that escape broadly neutralizing antibodies have been reported2,3. The identification of broadly neutralizing antibody classes that can neutralize viral escape mutants is critical for universal influenza virus vaccine design. Here we report a distinct class of broadly neutralizing antibodies that target a discrete membrane-proximal anchor epitope of the haemagglutinin stalk domain. Anchor epitope-targeting antibodies are broadly neutralizing across H1 viruses and can cross-react with H2 and H5 viruses that are a pandemic threat. Antibodies that target this anchor epitope utilize a highly restricted repertoire, which encodes two public binding motifs that make extensive contacts with conserved residues in the fusion peptide. Moreover, anchor epitope-targeting B cells are common in the human memory B cell repertoire and were recalled in humans by an oil-in-water adjuvanted chimeric haemagglutinin vaccine4,5, which is a potential universal influenza virus vaccine. To maximize protection against seasonal and pandemic influenza viruses, vaccines should aim to boost this previously untapped source of broadly neutralizing antibodies that are widespread in the human memory B cell pool.
© 2021. The Author(s).
Conflict of interest statement
The Icahn School of Medicine at Mount Sinai has patents (10736956, 10583188, 10137189, 10131695, 9968670, 9371366) and has submitted patent applications (10736956, 10583188, 10137189, 10131695, 9968670, 9371366) on universal influenza virus vaccines naming A.G.-S., R.N., P.P. and F.K. as inventors. The University of Chicago has submitted patent applications (632292804) on anti-anchor mAbs naming J.J.G. and P.C.W. as inventors. The A.G.-S. laboratory has received research support from Pfizer, Senhwa Biosciences, Kenall Manufacturing, Avimex, Johnson & Johnson, Dynavax, 7Hills Pharma, Pharmamar, ImmunityBio, Accurius, Nanocomposix, Hexamer, N-fold LLC and Merck, outside of the reported work. A.G.-S. has consulting agreements for the following companies involving cash and/or stock: Vivaldi Biosciences, Contrafect, 7Hills Pharma, Avimex, Vaxalto, Pagoda, Accurius, Esperovax, Farmak, Applied Biological Laboratories and Pfizer, outside of the reported work.
Figures
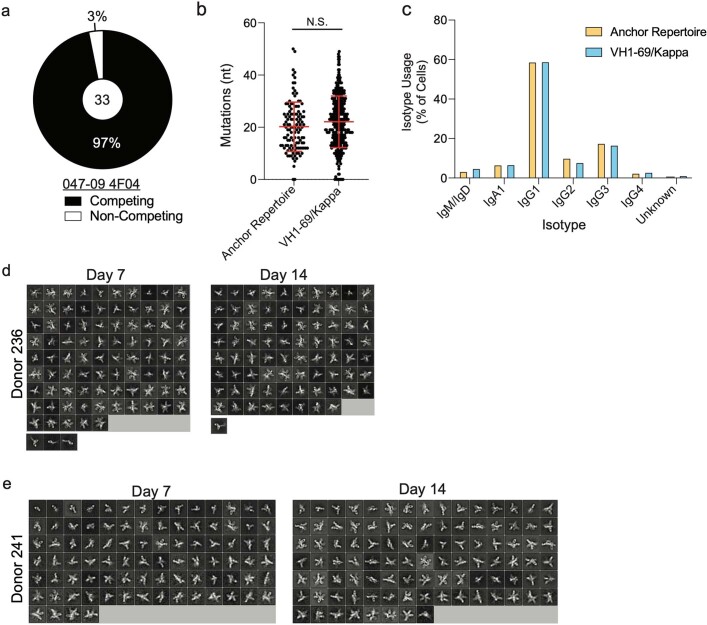
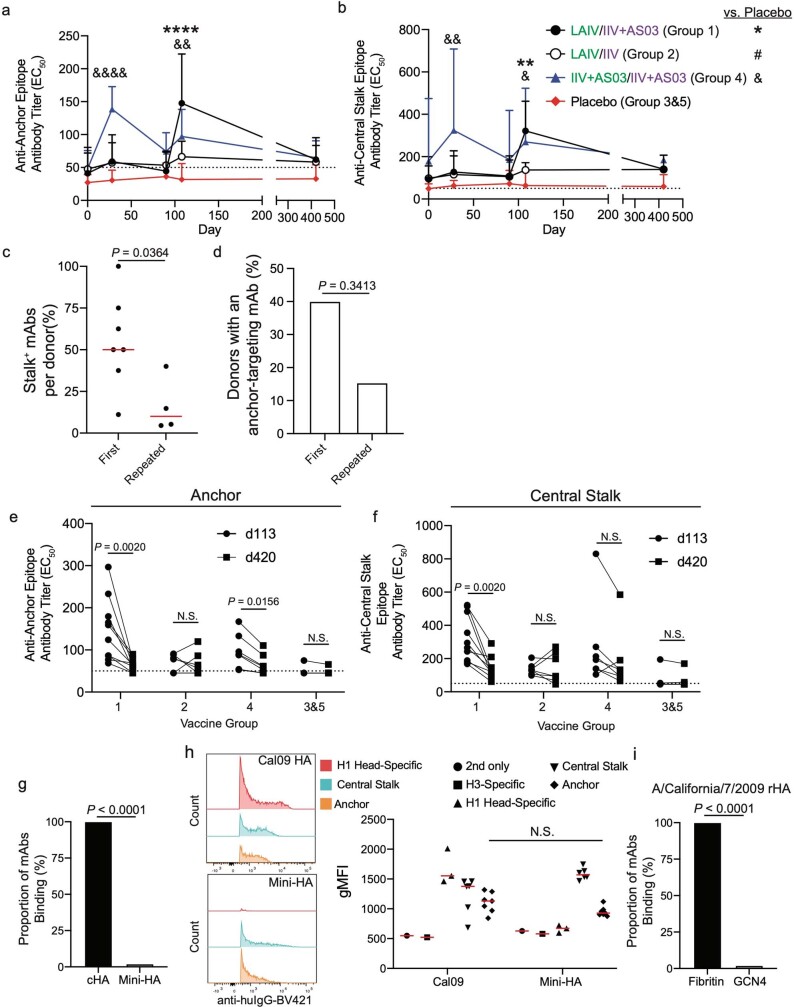
References
-
- Paules CI, Marston HD, Eisinger RW, Baltimore D, Fauci AS. The pathway to a universal influenza vaccine. Immunity. 2017;47:599–603. - PubMed
-
- Nachbagauer R, et al. A chimeric hemagglutinin-based universal influenza virus vaccine approach induces broad and long-lasting immunity in a randomized, placebo-controlled phase I trial. Nat. Med. 2020;27:106–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
