

The Functions of β-Defensin in Flounder (Paralichthys olivaceus): Antibiosis, Chemotaxis and Modulation of Phagocytosis
- PMID: 34943162
- PMCID: PMC8698591
- DOI: 10.3390/biology10121247
The Functions of β-Defensin in Flounder (Paralichthys olivaceus): Antibiosis, Chemotaxis and Modulation of Phagocytosis
Abstract
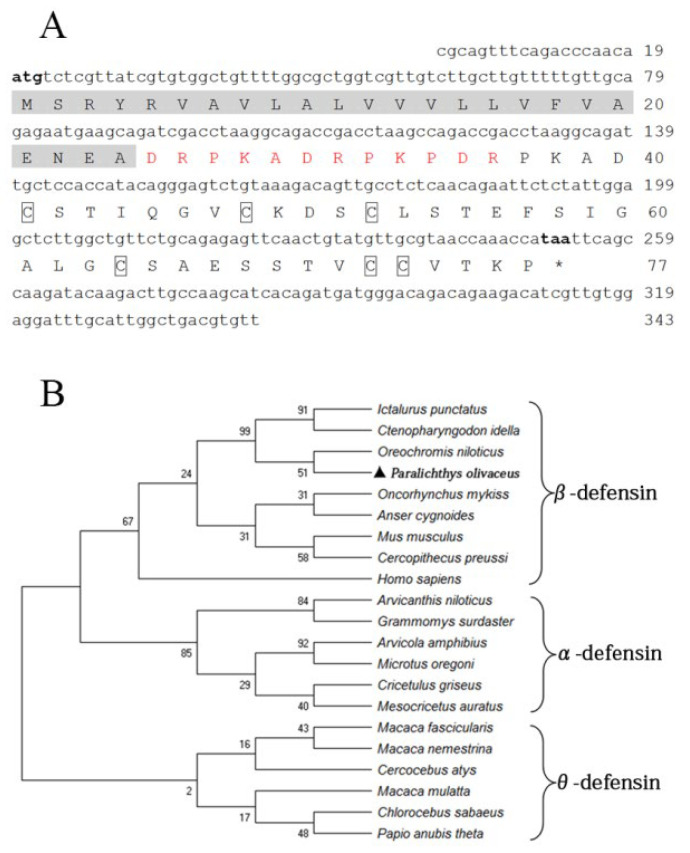
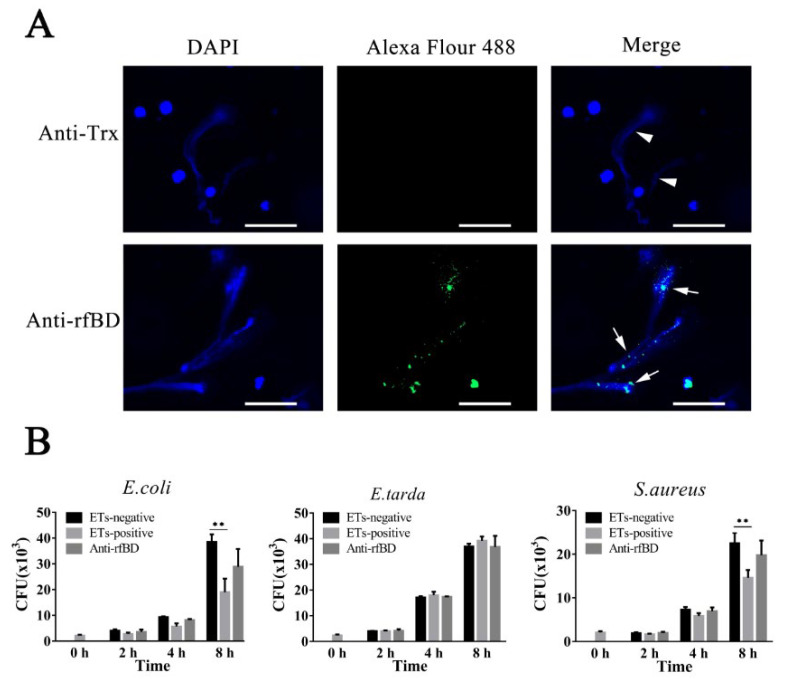
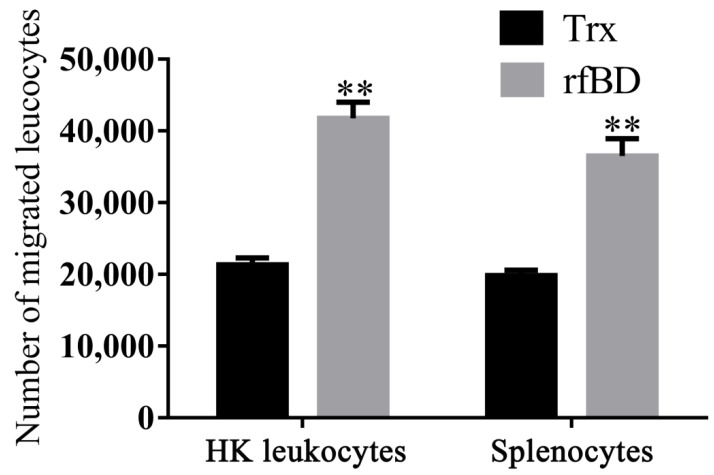
Most defensins are cationic antimicrobial peptides with broad-spectrum killing activity against bacteria, fungi and enveloped viruses. However, it should be recognized that there are some non-cationic β-defensins in organisms, which need to be further studied. In this study, a new spliced isoform of anionic β-defensin from flounder (Paralichthys olivaceus, fBD) was identified, and its antibiosis, chemotaxis and modulation of phagocytosis were examined. In addition, the contributions of fBD to the antimicrobial activity of extracellular traps (ETs) were also analyzed. The recombinant fBD (rfBD) could effectively inhibit the growth of Gram-positive bacteria (S. aureus, Micrococcus luteus) and Gram-negative bacteria (E. coli, V. alginolyticus, V. anguillarum). An indirect immunofluorescence assay showed that the fBD was co-localized in the extracellular traps released by the leukocytes. When the ETs were blocked with antibodies against rfBD, the proliferation of S. aureus and E. coli incubated with ETs tended to increase compared with that in the control group. In addition, the results obtained by flow cytometry showed that the rfBD could significantly chemoattract leukocytes and increase phagocytic activity in vitro. In conclusion, this study provides new insights into the biological function of anionic defensins, which can serve as one of the important effectors in extracellular traps and as a bridge between innate and adaptive immunity in teleosts.
Keywords: antibiosis; chemotaxis; extracellular traps; phagocytosis; β-defensin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Grants and funding
LinkOut - more resources
Full Text Sources
