

Exosomes originating from infection with the cytoplasmic single-stranded RNA virus Rift Valley fever virus (RVFV) protect recipient cells by inducing RIG-I mediated IFN-B response that leads to activation of autophagy
- PMID: 34953502
- PMCID: PMC8710069
- DOI: 10.1186/s13578-021-00732-z
Exosomes originating from infection with the cytoplasmic single-stranded RNA virus Rift Valley fever virus (RVFV) protect recipient cells by inducing RIG-I mediated IFN-B response that leads to activation of autophagy
Abstract
Background: Although multiple studies have demonstrated a role for exosomes during virus infections, our understanding of the mechanisms by which exosome exchange regulates immune response during viral infections and affects viral pathogenesis is still in its infancy. In particular, very little is known for cytoplasmic single-stranded RNA viruses such as SARS-CoV-2 and Rift Valley fever virus (RVFV). We have used RVFV infection as a model for cytoplasmic single-stranded RNA viruses to address this gap in knowledge. RVFV is a highly pathogenic agent that causes RVF, a zoonotic disease for which no effective therapeutic or approved human vaccine exist.
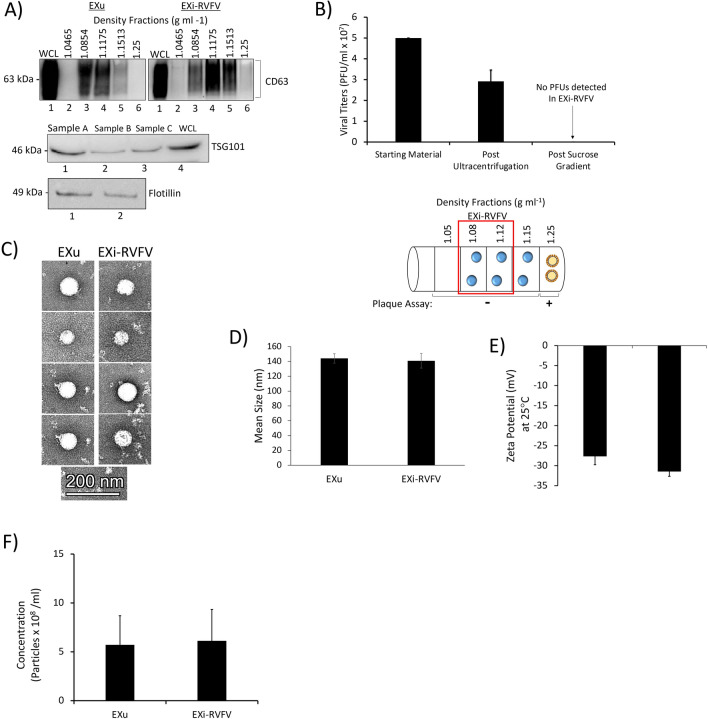
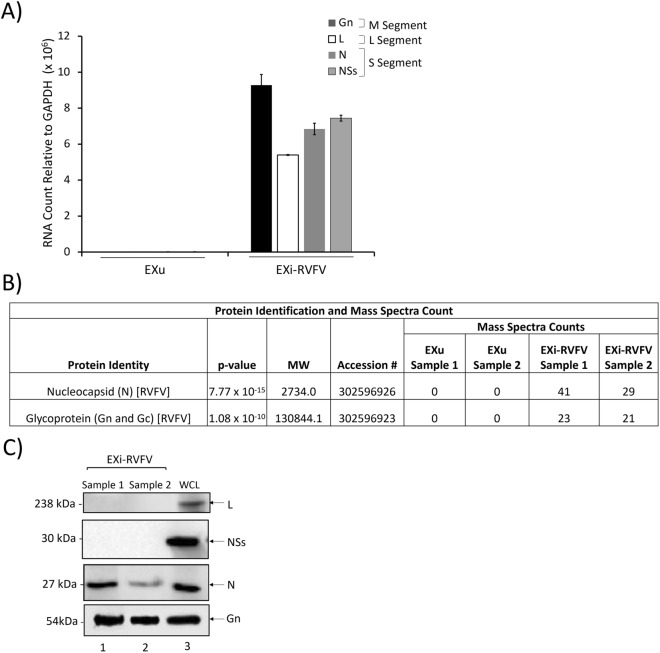
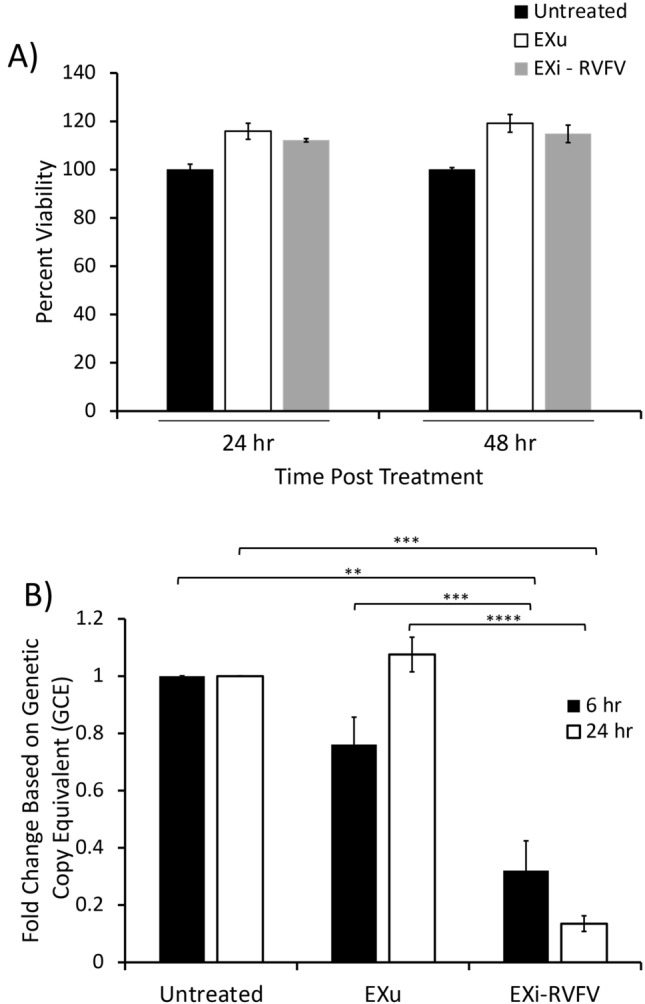
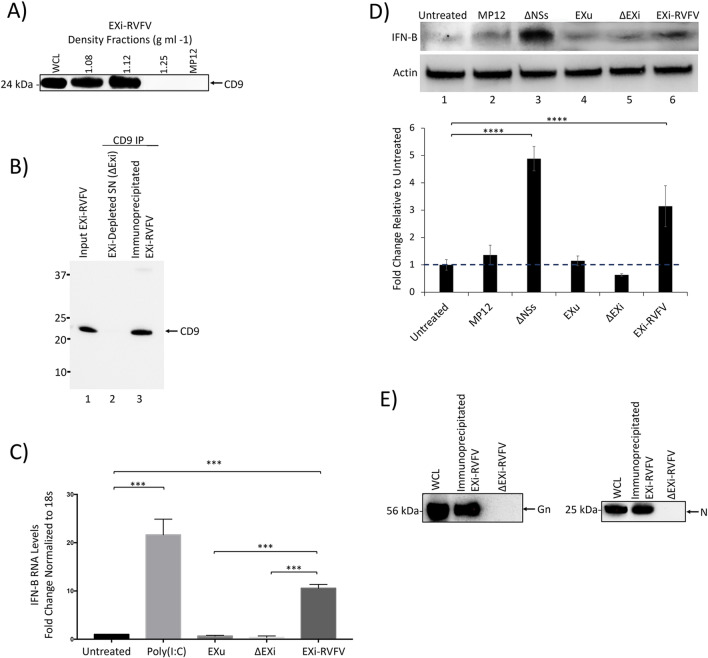
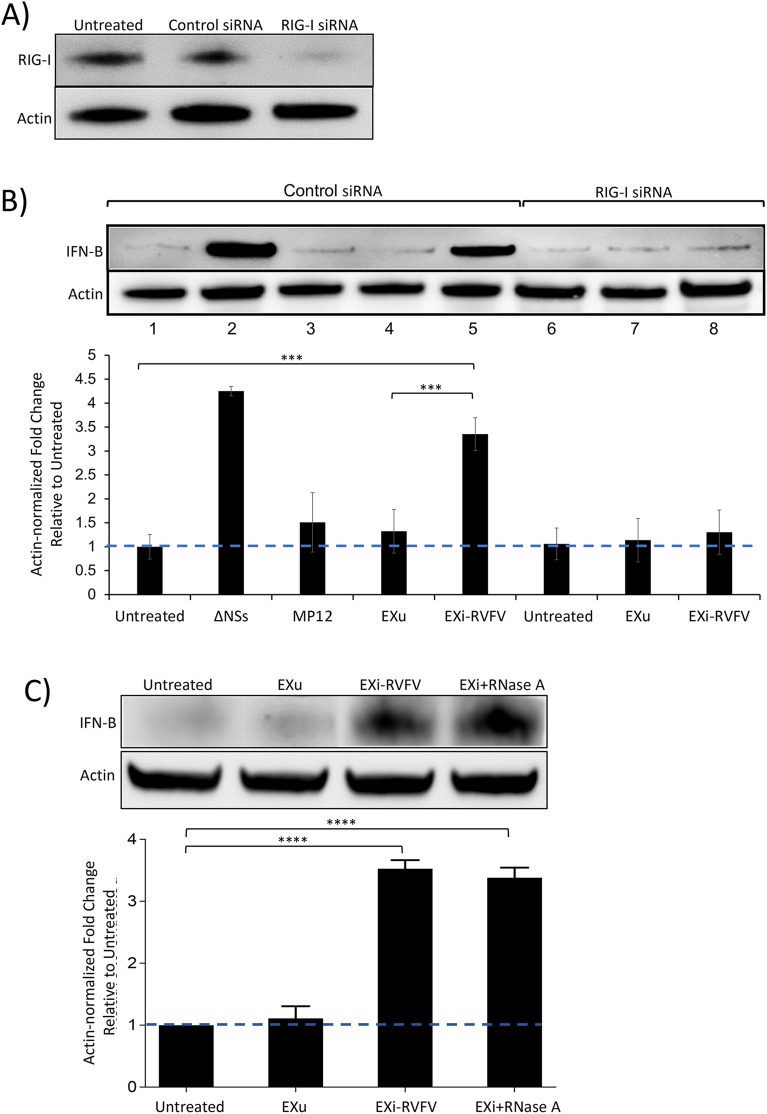
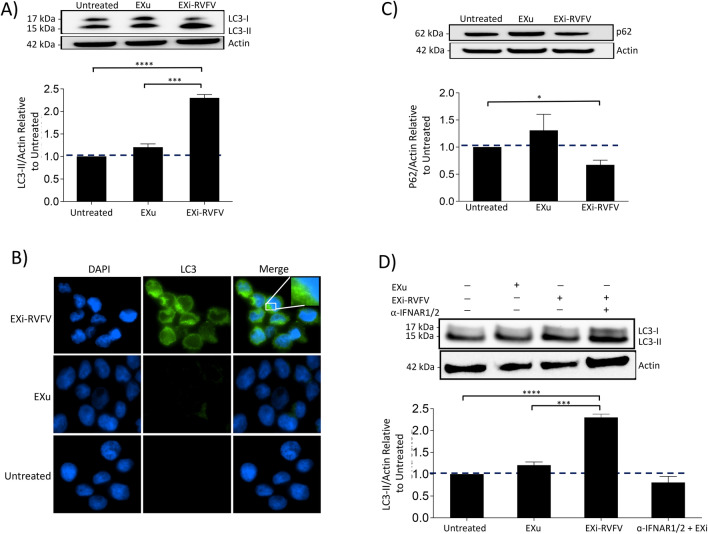
Results: We show here that exosomes released from cells infected with RVFV (designated as EXi-RVFV) serve a protective role for the host and provide a mechanistic model for these effects. Our results show that treatment of both naïve immune cells (U937 monocytes) and naïve non-immune cells (HSAECs) with EXi-RVFV induces a strong RIG-I dependent activation of IFN-B. We also demonstrate that this strong anti-viral response leads to activation of autophagy in treated cells and correlates with resistance to subsequent viral infection. Since we have shown that viral RNA genome is associated with EXi-RVFV, RIG-I activation might be mediated by the presence of packaged viral RNA sequences.
Conclusions: Using RVFV infection as a model for cytoplasmic single-stranded RNA viruses, our results show a novel mechanism of host protection by exosomes released from infected cells (EXi) whereby the EXi activate RIG-I to induce IFN-dependent activation of autophagy in naïve recipient cells including monocytes. Because monocytes serve as reservoirs for RVFV replication, this EXi-RVFV-induced activation of autophagy in monocytes may work to slow down or halt viral dissemination in the infected organism. These findings offer novel mechanistic insights that may aid in future development of effective vaccines or therapeutics, and that may be applicable for a better molecular understanding of how exosome release regulates innate immune response to other cytoplasmic single-stranded RNA viruses.
Keywords: Autophagy; Exosome; IFN-B; Innate immune response; RIG-I; Rift Valley fever virus; SARS-CoV-2; Single-stranded RNA virus; Viral RNA; Virus infection.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
