

The Nucleocapsid of Paramyxoviruses: Structure and Function of an Encapsidated Template
- PMID: 34960734
- PMCID: PMC8708338
- DOI: 10.3390/v13122465
The Nucleocapsid of Paramyxoviruses: Structure and Function of an Encapsidated Template
Abstract
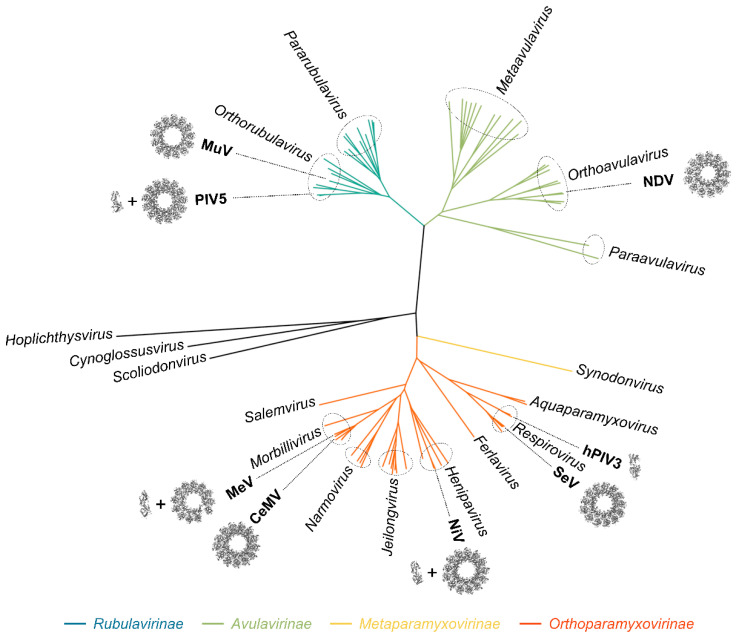
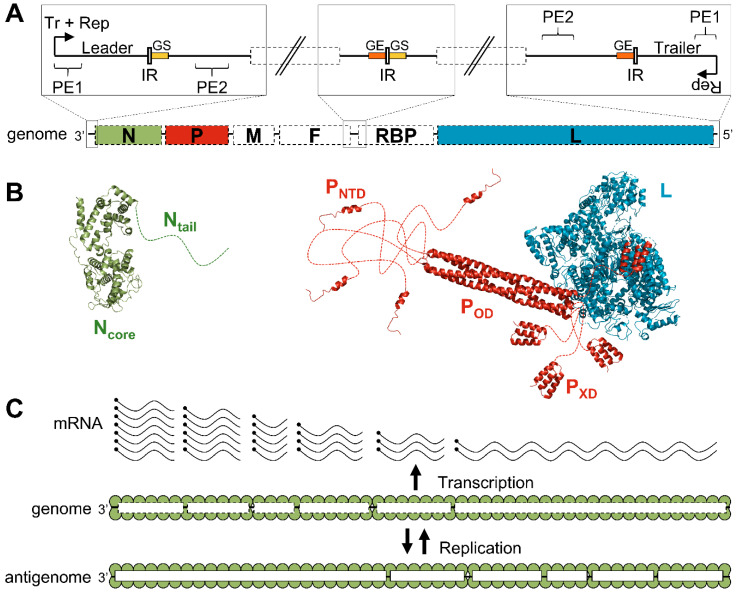
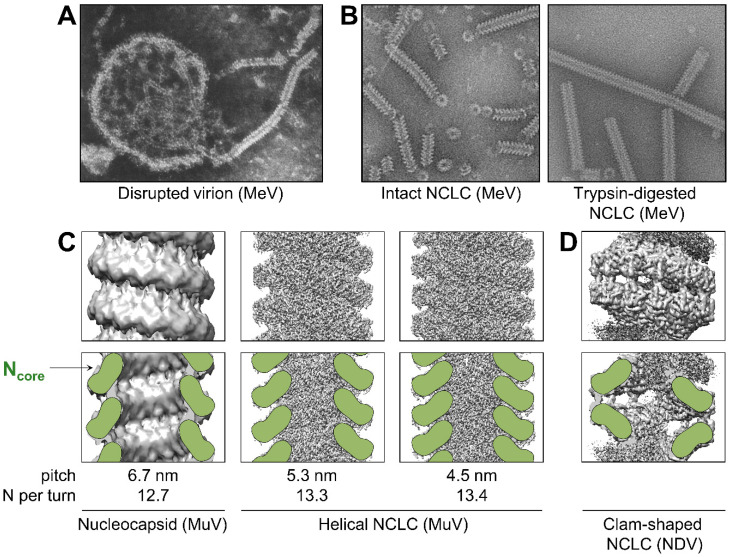
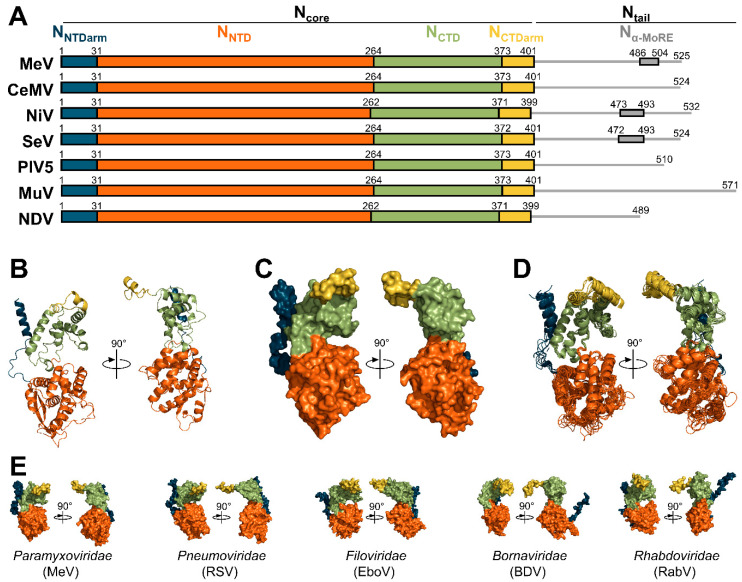
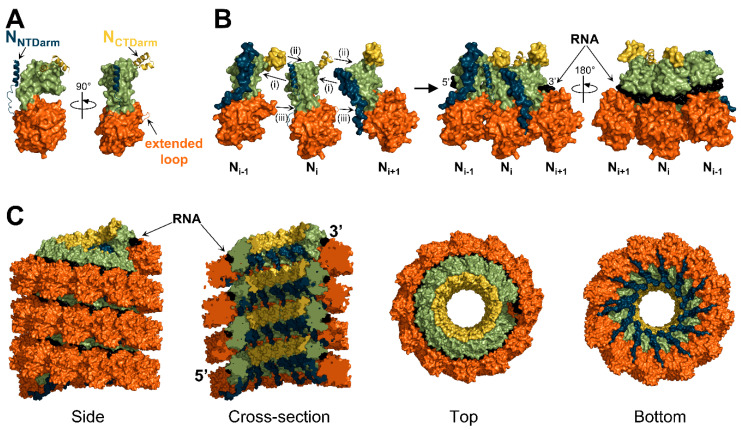
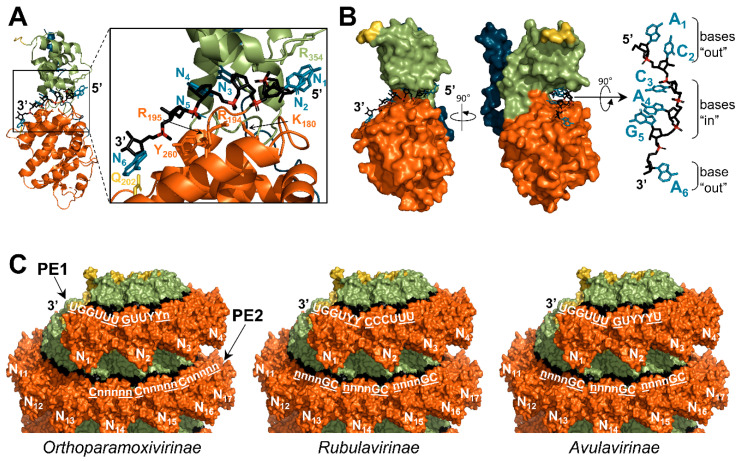
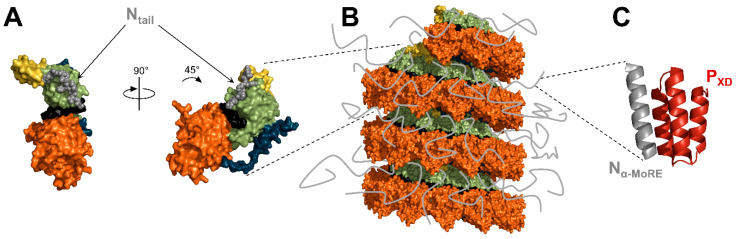
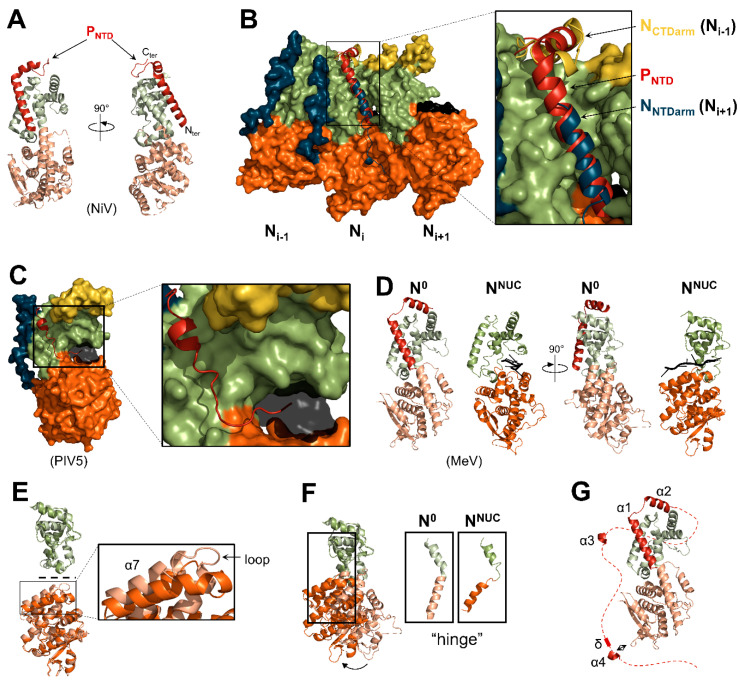
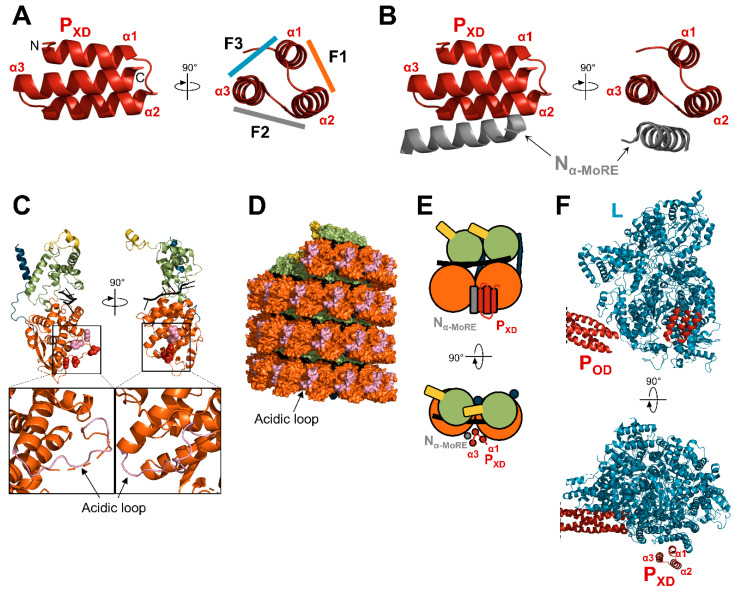
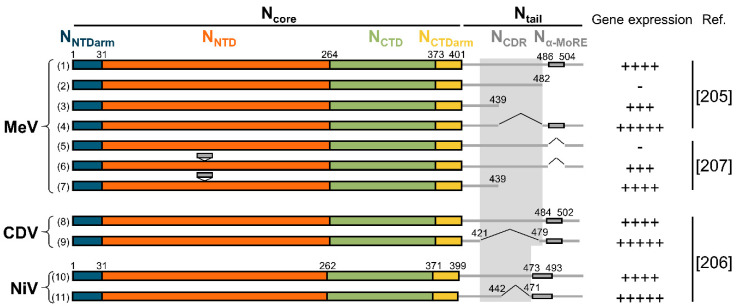
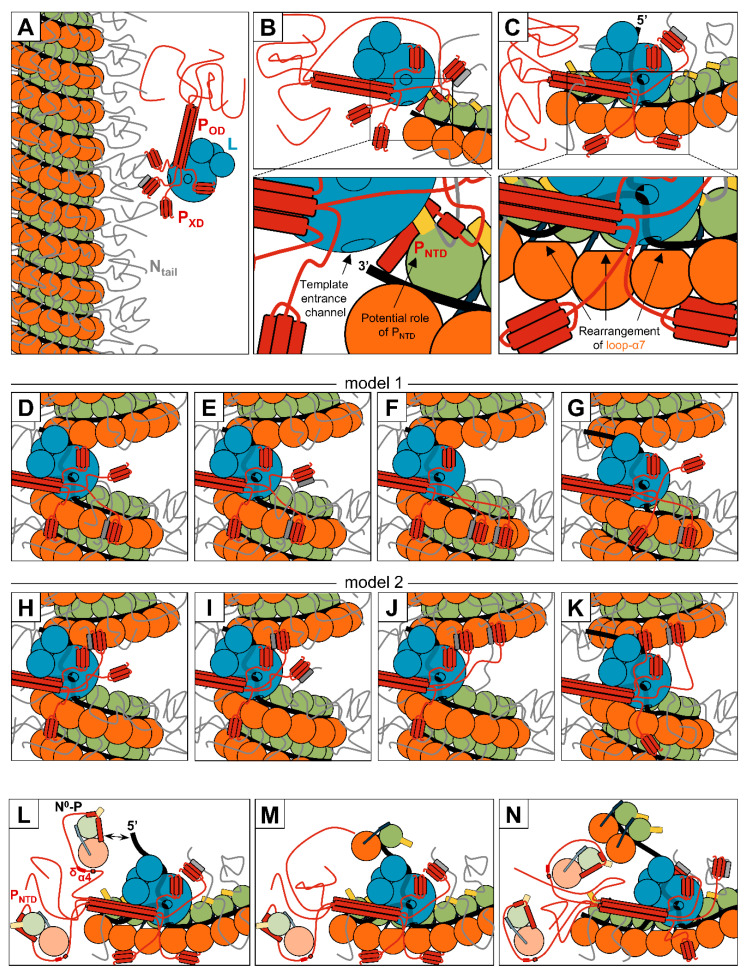
Viruses of the Paramyxoviridae family share a common and complex molecular machinery for transcribing and replicating their genomes. Their non-segmented, negative-strand RNA genome is encased in a tight homopolymer of viral nucleoproteins (N). This ribonucleoprotein complex, termed a nucleocapsid, is the template of the viral polymerase complex made of the large protein (L) and its co-factor, the phosphoprotein (P). This review summarizes the current knowledge on several aspects of paramyxovirus transcription and replication, including structural and functional data on (1) the architecture of the nucleocapsid (structure of the nucleoprotein, interprotomer contacts, interaction with RNA, and organization of the disordered C-terminal tail of N), (2) the encapsidation of the genomic RNAs (structure of the nucleoprotein in complex with its chaperon P and kinetics of RNA encapsidation in vitro), and (3) the use of the nucleocapsid as a template for the polymerase complex (release of the encased RNA and interaction network allowing the progress of the polymerase complex). Finally, this review presents models of paramyxovirus transcription and replication.
Keywords: intrinsic disorder; nucleocapsid; nucleoprotein; paramyxoviruses; phosphoprotein; polymerase complex.
Conflict of interest statement
The author declares no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
