

Here comes the sun: How optimization of photosynthetic light reactions can boost crop yields
- PMID: 34962073
- PMCID: PMC9302994
- DOI: 10.1111/jipb.13206
Here comes the sun: How optimization of photosynthetic light reactions can boost crop yields
Abstract
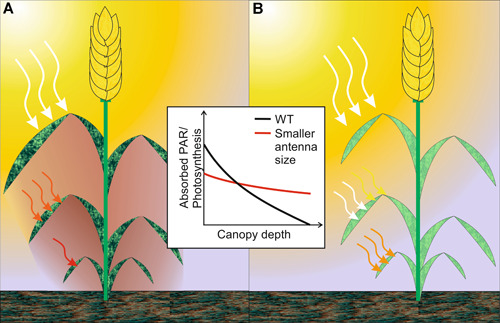
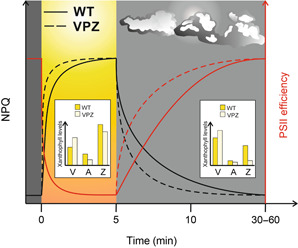
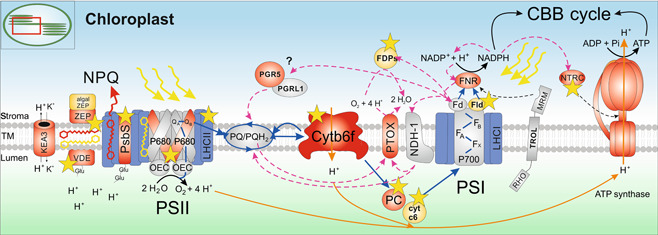
Photosynthesis started to evolve some 3.5 billion years ago CO2 is the substrate for photosynthesis and in the past 200-250 years, atmospheric levels have approximately doubled due to human industrial activities. However, this time span is not sufficient for adaptation mechanisms of photosynthesis to be evolutionarily manifested. Steep increases in human population, shortage of arable land and food, and climate change call for actions, now. Thanks to substantial research efforts and advances in the last century, basic knowledge of photosynthetic and primary metabolic processes can now be translated into strategies to optimize photosynthesis to its full potential in order to improve crop yields and food supply for the future. Many different approaches have been proposed in recent years, some of which have already proven successful in different crop species. Here, we summarize recent advances on modifications of the complex network of photosynthetic light reactions. These are the starting point of all biomass production and supply the energy equivalents necessary for downstream processes as well as the oxygen we breathe.
Keywords: bioengineering; crop improvement; electron transfer; light reactions; photosynthesis; photosystem; stress tolerance.
© 2022 The Authors. Journal of Integrative Plant Biology published by John Wiley & Sons Australia, Ltd on behalf of Institute of Botany, Chinese Academy of Sciences.
Conflict of interest statement
The authors declare there are no conflicts of interest.
Figures
References
-
- Abbasi, A.Z. , Bilal, M. , Khurshid, G. , Yiotis, C. , Zeb, I. , Hussain, J. , Baig, A. , Shah, M.M. , Chaudhary, S.U. , Osborne, B. , and Ahmad, R. (2021). Expression of cyanobacterial genes enhanced CO2 assimilation and biomass production in transgenic Arabidopsis thaliana . PeerJ 9: e11860. - PMC - PubMed
-
- Abdel‐Ghany, S.E. (2009). Contribution of plastocyanin isoforms to photosynthesis and homeostasis in Arabidopsis thaliana grown at different copper regimes. Planta 229: 767–779. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
