

Evolution of Olfactory Receptors Tuned to Mustard Oils in Herbivorous Drosophilidae
- PMID: 34963012
- PMCID: PMC8826531
- DOI: 10.1093/molbev/msab362
Evolution of Olfactory Receptors Tuned to Mustard Oils in Herbivorous Drosophilidae
Abstract
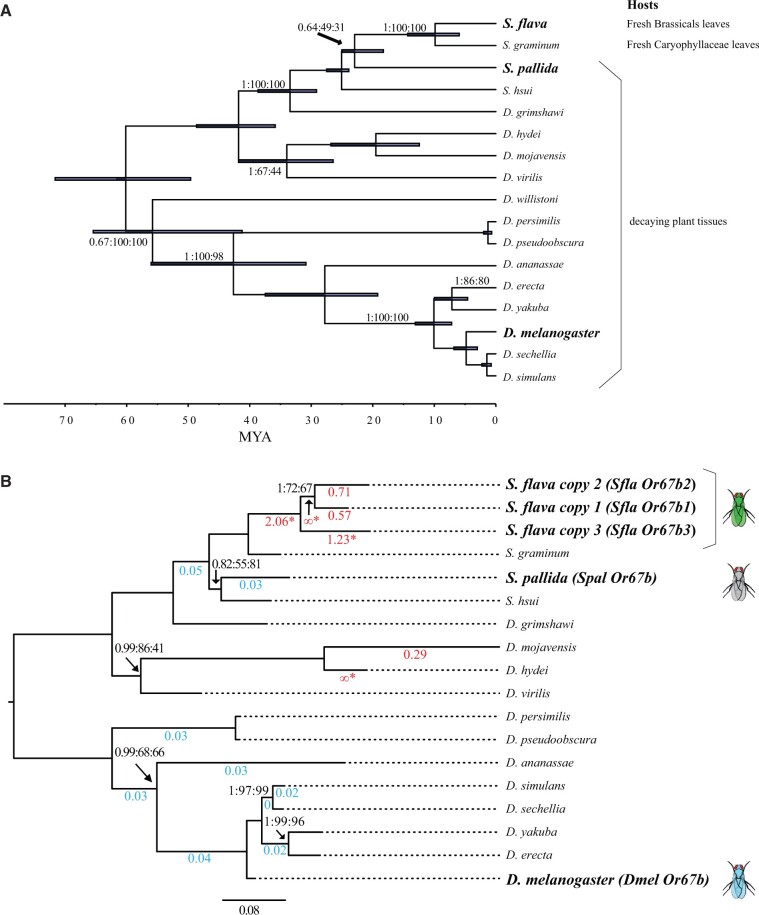
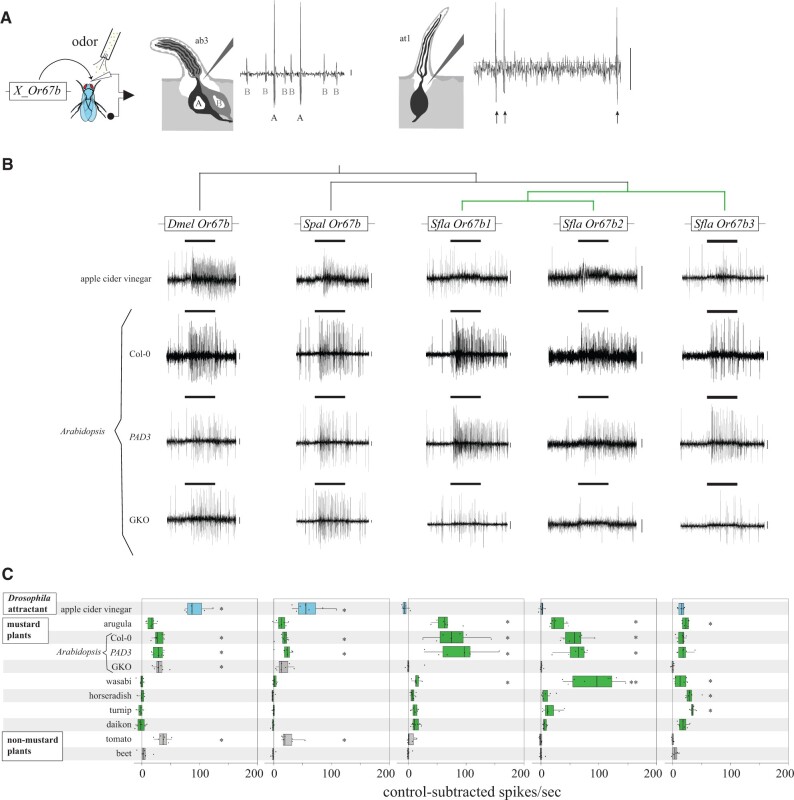
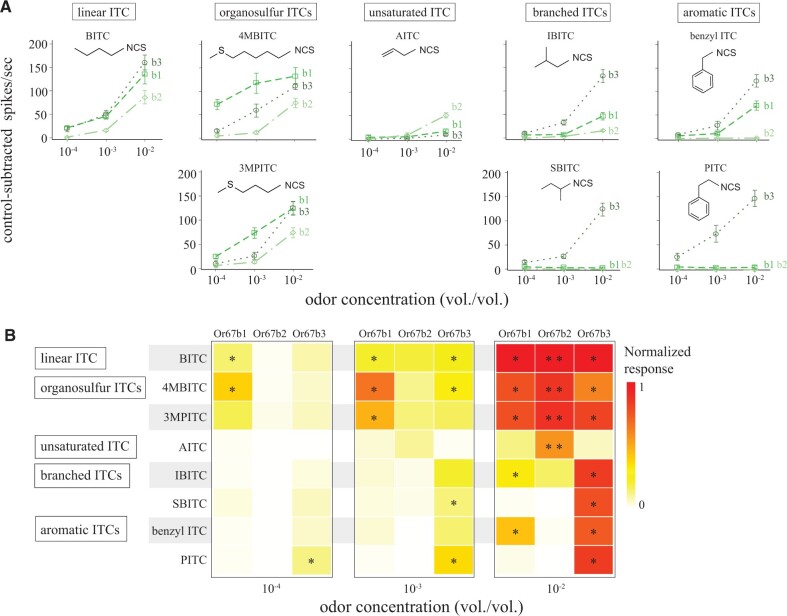
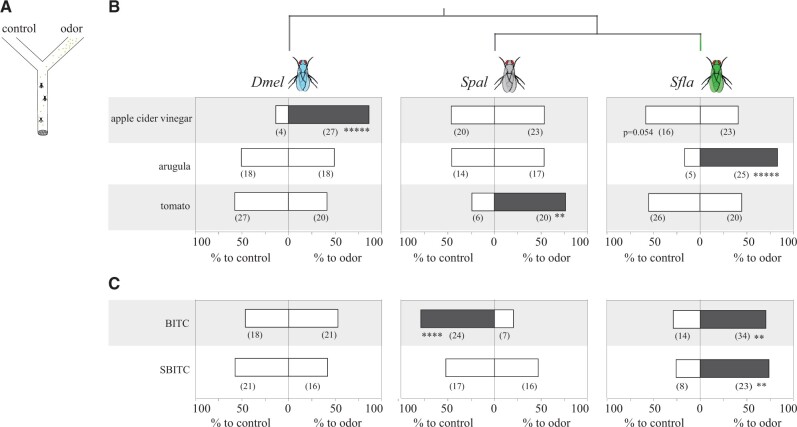
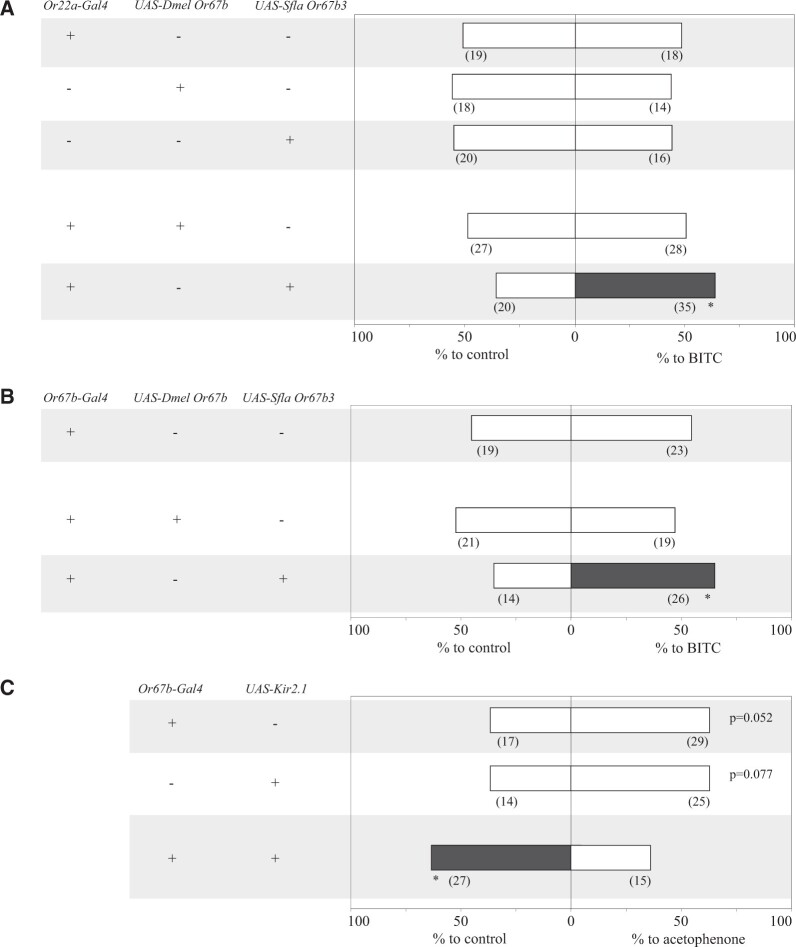
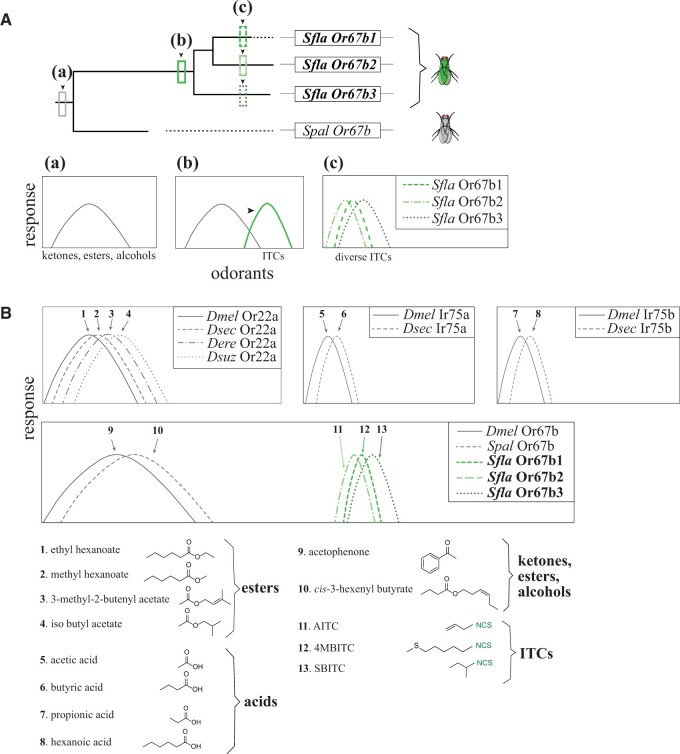
The diversity of herbivorous insects is attributed to their propensity to specialize on toxic plants. In an evolutionary twist, toxins betray the identity of their bearers when herbivores coopt them as cues for host-plant finding, but the evolutionary mechanisms underlying this phenomenon are poorly understood. We focused on Scaptomyza flava, an herbivorous drosophilid specialized on isothiocyanate (ITC)-producing (Brassicales) plants, and identified Or67b paralogs that were triplicated as mustard-specific herbivory evolved. Using in vivo heterologous systems for the expression of olfactory receptors, we found that S. flava Or67bs, but not the homologs from microbe-feeding relatives, responded selectively to ITCs, each paralog detecting different ITC subsets. Consistent with this, S. flava was attracted to ITCs, as was Drosophila melanogaster expressing S. flava Or67b3 in the homologous Or67b olfactory circuit. ITCs were likely coopted as olfactory attractants through gene duplication and functional specialization (neofunctionalization and subfunctionalization) in S. flava, a recently derived herbivore.
Keywords: Drosophila melanogaster; Scaptomyza flava; Brassicales; Or67b; SSR; chemoreceptor; evolution; gene duplication; herbivory; isothiocyanate; neofunctionalization; olfaction; olfactory receptor; olfactory specialization; specialization; subfunctionalization; wasabi.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures

References
-
- Aurand LW, Singleton JA, Bell TA, Etchells JL.. 1966. Volatile components in the vapors of natural and distilled vinegars. J Food Sci. 31(2):172–177.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
