

The imprinted Igf2-Igf2r axis is critical for matching placental microvasculature expansion to fetal growth
- PMID: 34963058
- PMCID: PMC8751640
- DOI: 10.1016/j.devcel.2021.12.005
The imprinted Igf2-Igf2r axis is critical for matching placental microvasculature expansion to fetal growth
Abstract
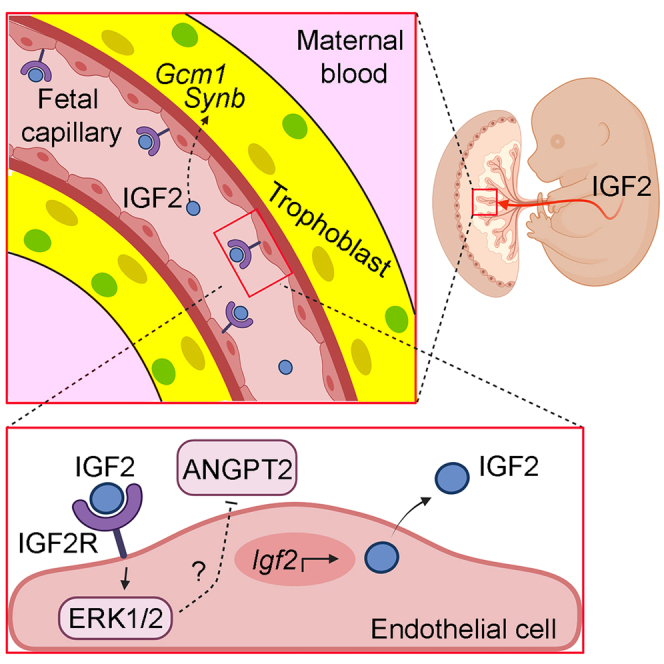
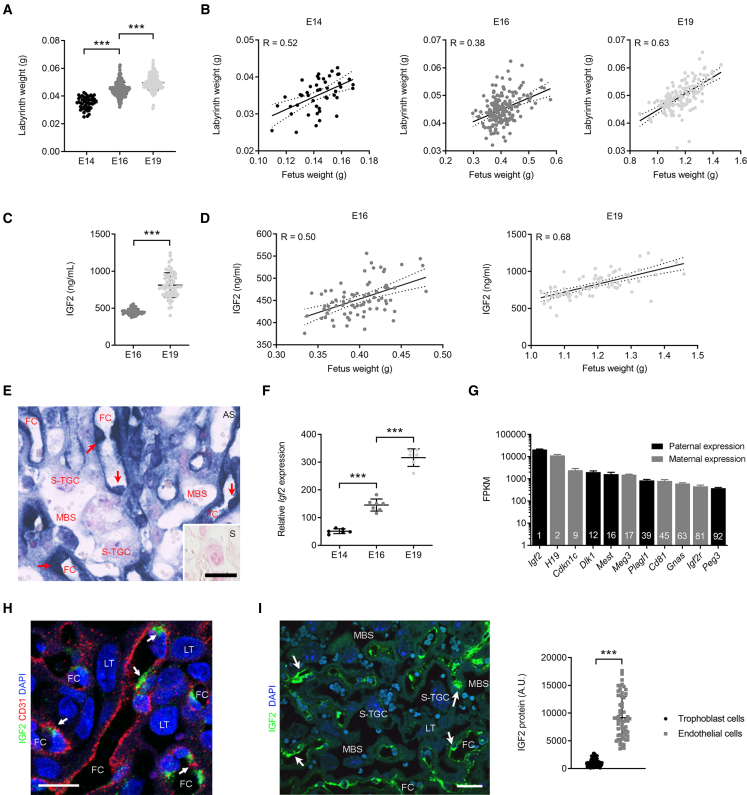
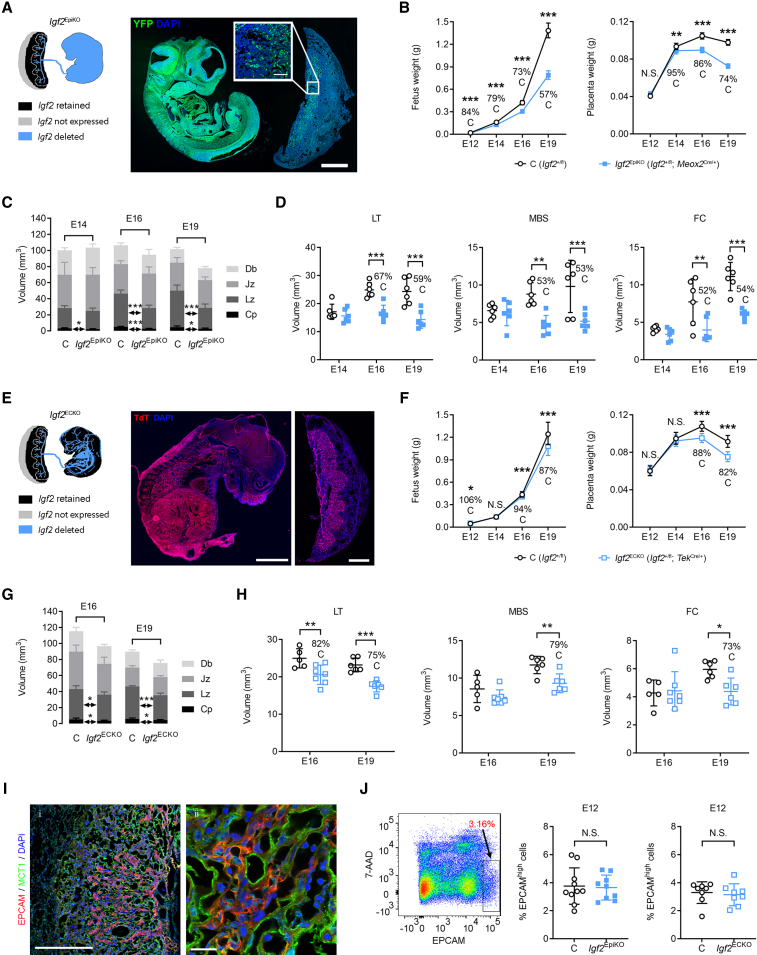
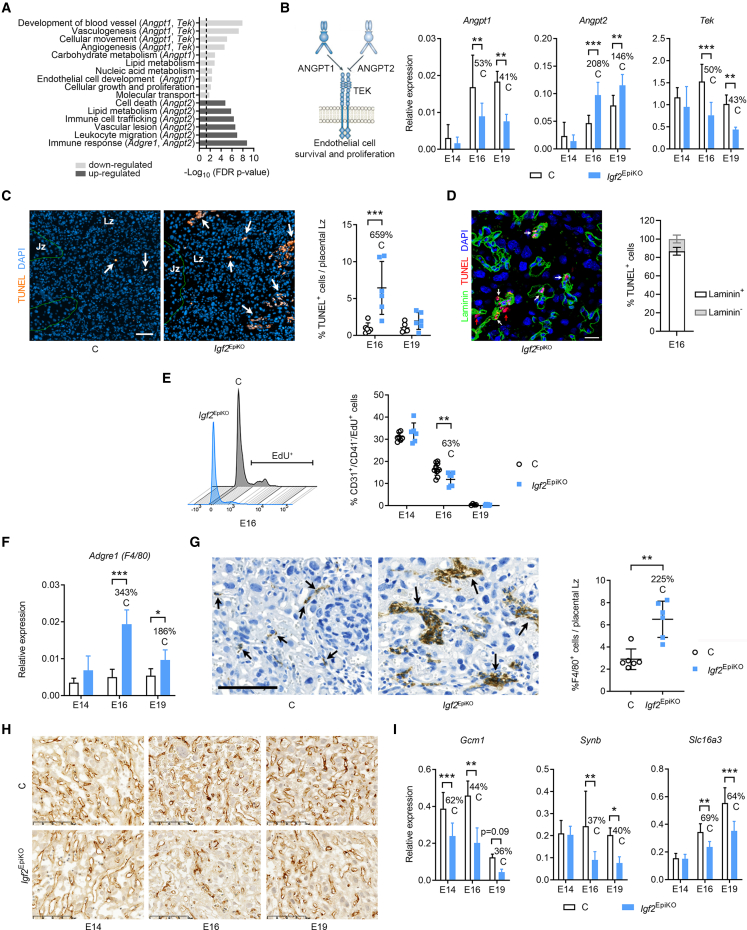
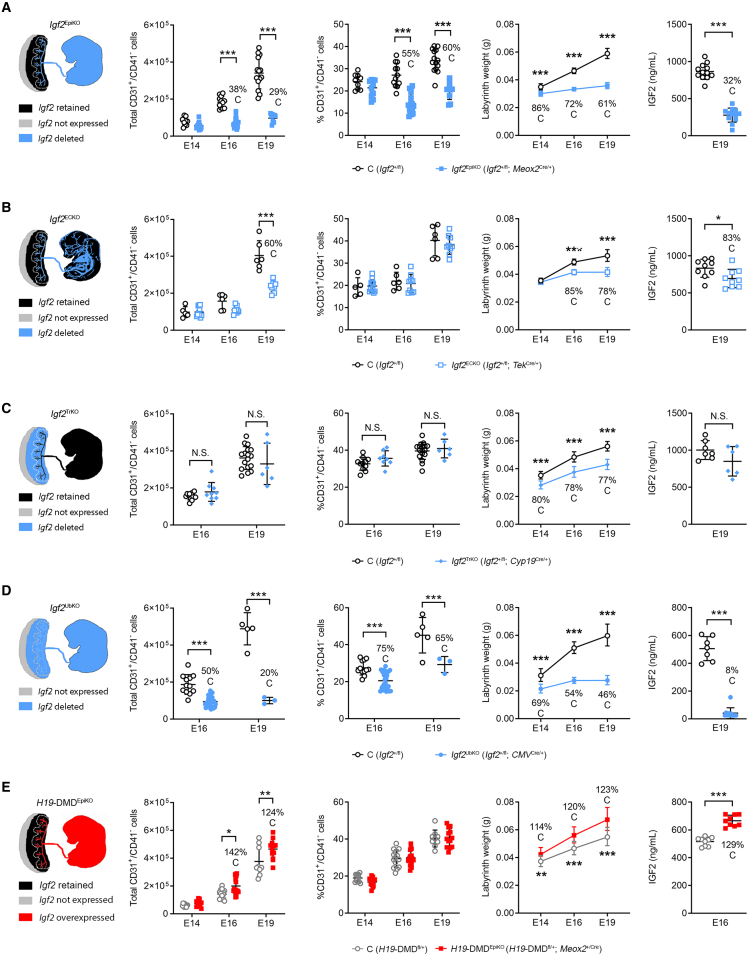
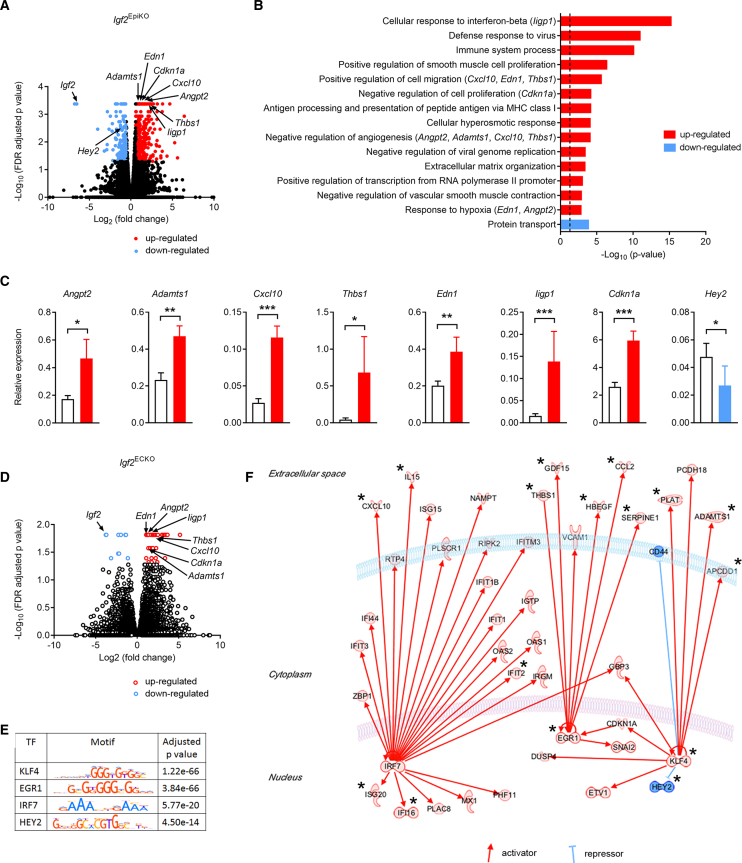
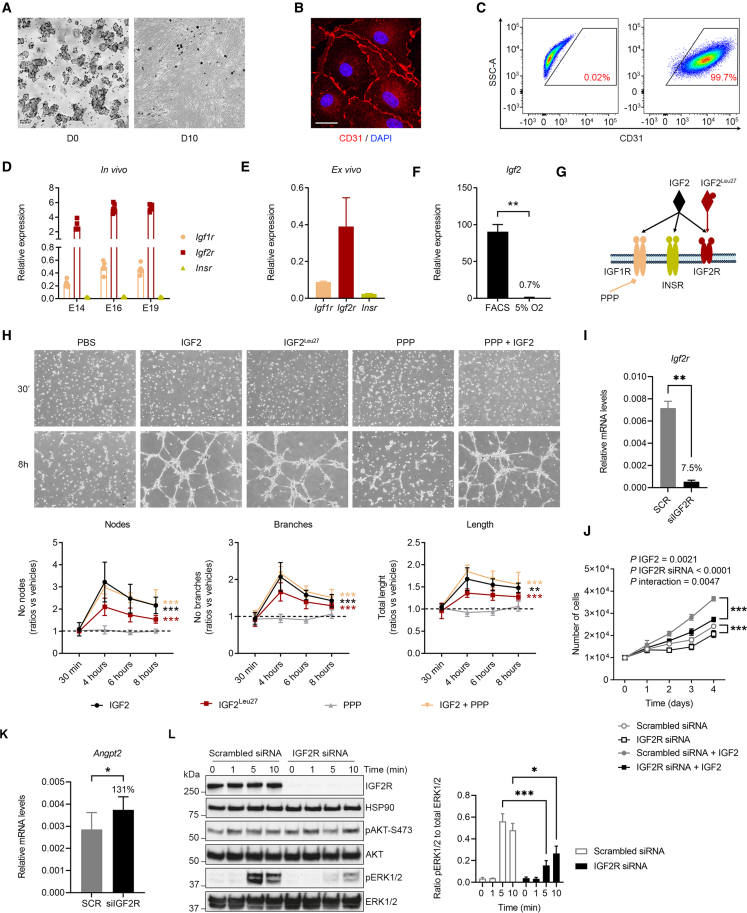
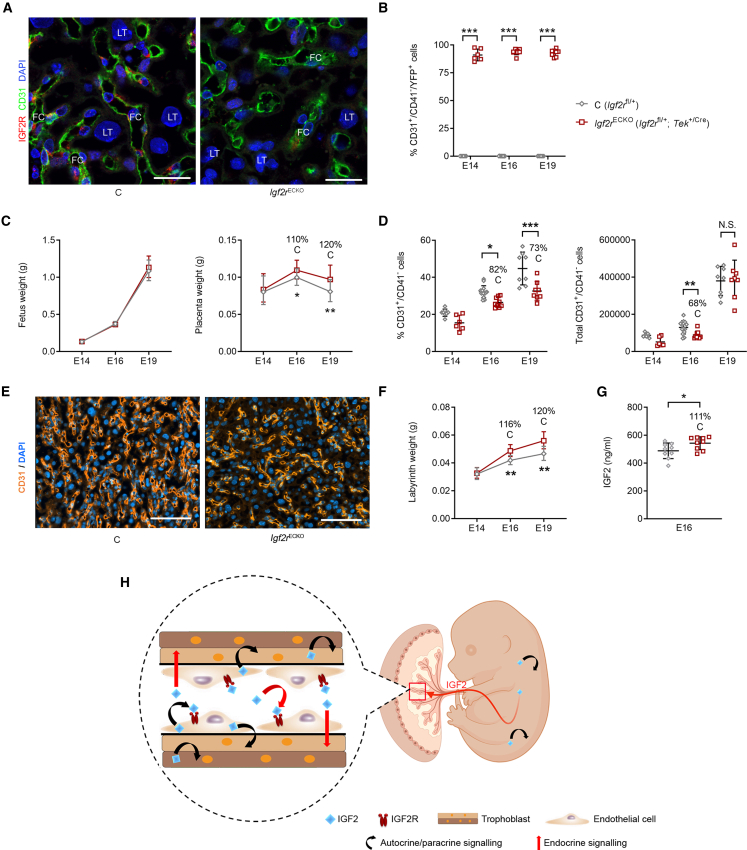
In all eutherian mammals, growth of the fetus is dependent upon a functional placenta, but whether and how the latter adapts to putative fetal signals is currently unknown. Here, we demonstrate, through fetal, endothelial, hematopoietic, and trophoblast-specific genetic manipulations in the mouse, that endothelial and fetus-derived IGF2 is required for the continuous expansion of the feto-placental microvasculature in late pregnancy. The angiocrine effects of IGF2 on placental microvasculature expansion are mediated, in part, through IGF2R and angiopoietin-Tie2/TEK signaling. Additionally, IGF2 exerts IGF2R-ERK1/2-dependent pro-proliferative and angiogenic effects on primary feto-placental endothelial cells ex vivo. Endothelial and fetus-derived IGF2 also plays an important role in trophoblast morphogenesis, acting through Gcm1 and Synb. Thus, our study reveals a direct role for the imprinted Igf2-Igf2r axis on matching placental development to fetal growth and establishes the principle that hormone-like signals from the fetus play important roles in controlling placental microvasculature and trophoblast morphogenesis.
Keywords: IGF2; IGF2R; angiogenesis; angiopoietins; development; endothelial cells; fetal growth; genomic imprinting; placenta; trophoblast morphogenesis.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Angiolini E., Coan P.M., Sandovici I., Iwajomo O.H., Peck G., Burton G.J., Sibley C.P., Reik W., Fowden A.L., Constância M. Developmental adaptations to increased fetal nutrient demand in mouse genetic models of Igf2-mediated overgrowth. FASEB J. 2011;25:1737–1745. doi: 10.1096/fj.10-175273. - DOI - PubMed
-
- Aoki A., Shiozaki A., Sameshima A., Higashimoto K., Soejima H., Saito S. Beckwith-Wiedemann syndrome with placental chorangioma due to H19-differentially methylated region hypermethylation: a case report. J. Obstet. Gynaecol. Res. 2011;37:1872–1876. doi: 10.1111/j.1447-0756.2011.01654.x. - DOI - PubMed
-
- Armes J.E., McGown I., Williams M., Broomfield A., Gough K., Lehane F., Lourie R. The placenta in Beckwith-Wiedemann syndrome: genotype-phenotype associations, excessive extravillous trophoblast and placental mesenchymal dysplasia. Pathology. 2012;44:519–527. doi: 10.1097/PAT.0b013e3283559c94. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MC_UU_12012/1/MRC_/Medical Research Council/United Kingdom
- MR/R022690/1/MRC_/Medical Research Council/United Kingdom
- BB/H003312/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MC_UU_00014/5/MRC_/Medical Research Council/United Kingdom
- DH_/Department of Health/United Kingdom
- MRC_MC_UU_12012/4/MRC_/Medical Research Council/United Kingdom
- MRC_MC_UU_12012/5/MRC_/Medical Research Council/United Kingdom
- MC_UU_00014/1/MRC_/Medical Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- MC_UU_00014/4/MRC_/Medical Research Council/United Kingdom
- MC_UU_12012/4/MRC_/Medical Research Council/United Kingdom
- MC_UU_12012/5/MRC_/Medical Research Council/United Kingdom
- 220456/Z/20/Z/WT_/Wellcome Trust/United Kingdom
- MR/S026193/1/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
