

Post-synaptic scaffold protein TANC2 in psychiatric and somatic disease risk
- PMID: 34964047
- PMCID: PMC8906171
- DOI: 10.1242/dmm.049205
Post-synaptic scaffold protein TANC2 in psychiatric and somatic disease risk
Abstract
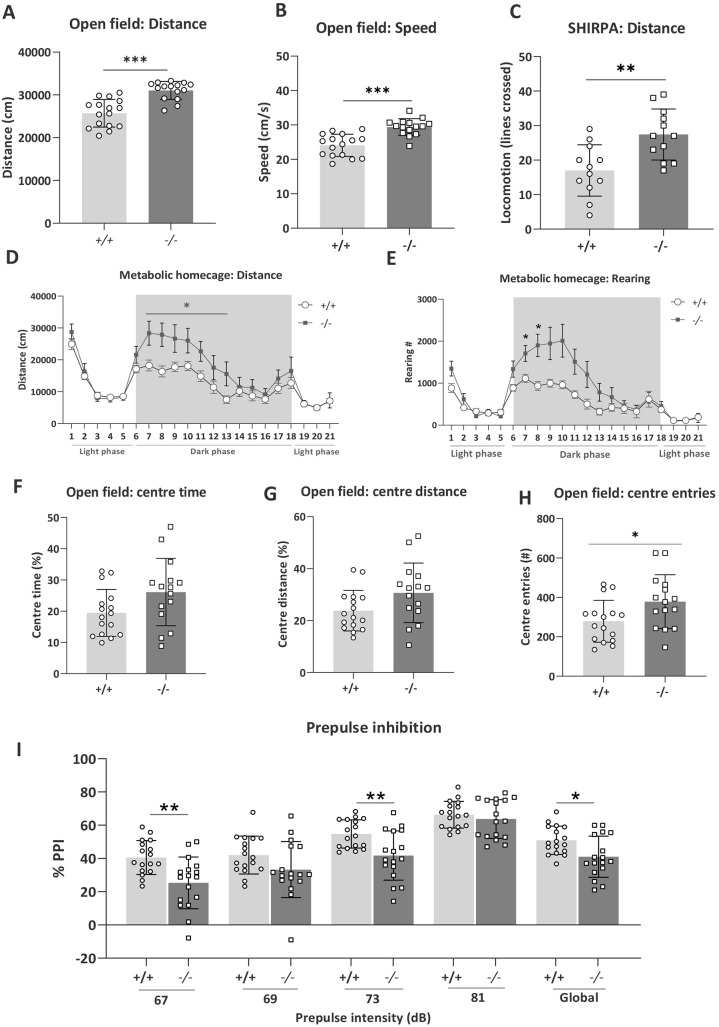
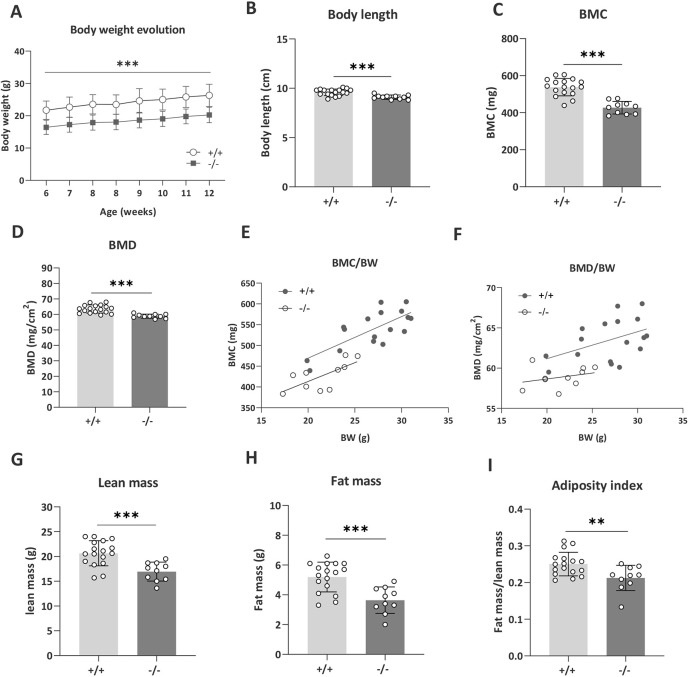
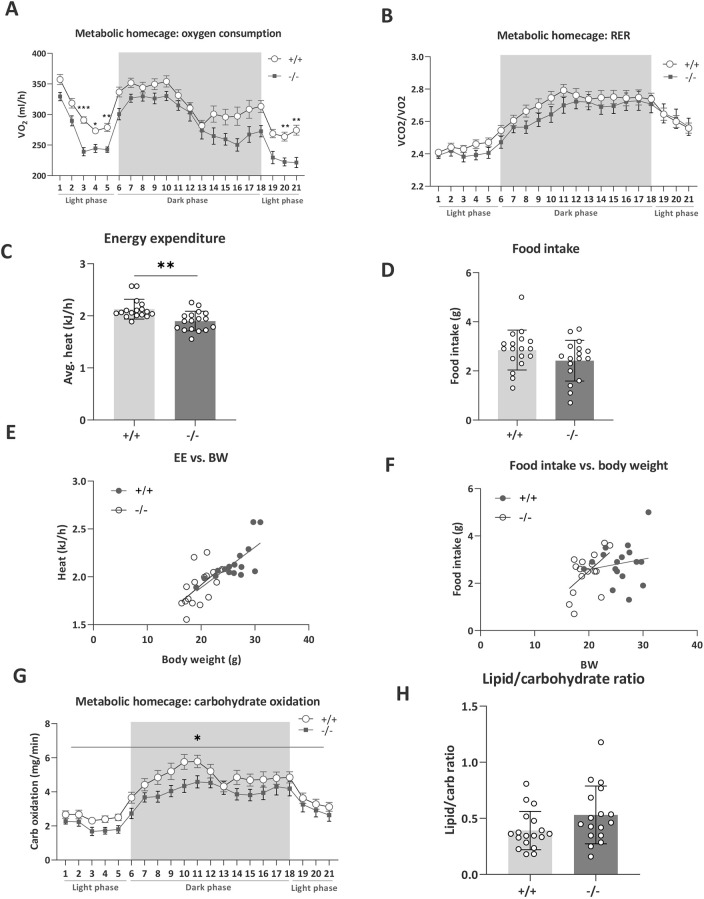
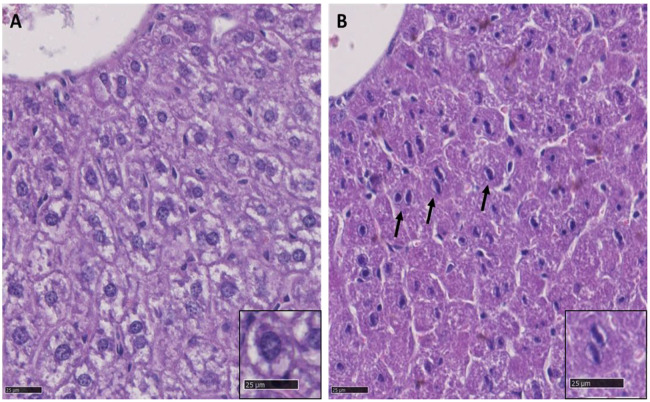
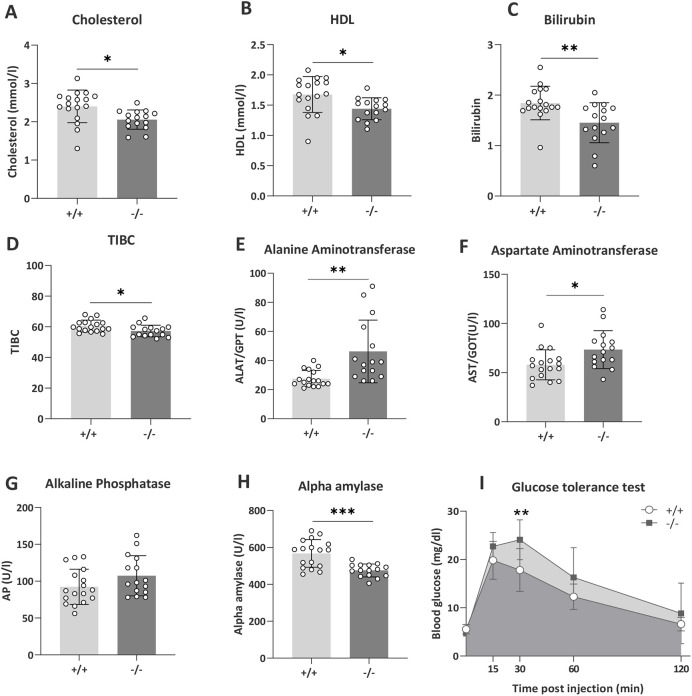
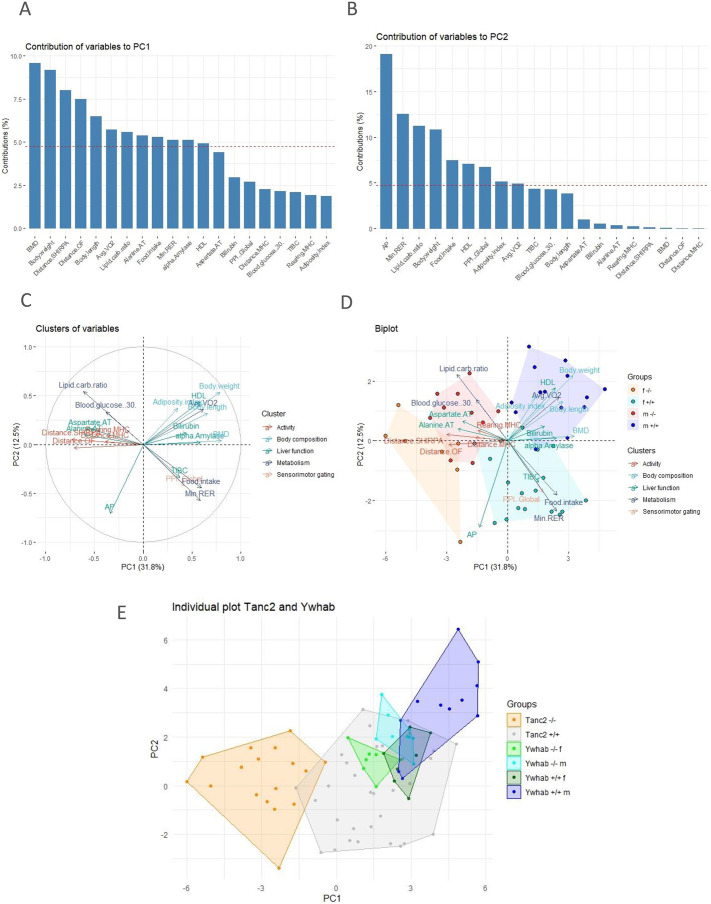
Understanding the shared genetic aetiology of psychiatric and medical comorbidity in neurodevelopmental disorders (NDDs) could improve patient diagnosis, stratification and treatment options. Rare tetratricopeptide repeat, ankyrin repeat and coiled-coil containing 2 (TANC2)-disrupting variants were disease causing in NDD patients. The post-synaptic scaffold protein TANC2 is essential for dendrite formation in synaptic plasticity and plays an unclarified but critical role in development. We here report a novel homozygous-viable Tanc2-disrupted function model in which mutant mice were hyperactive and had impaired sensorimotor gating consistent with NDD patient psychiatric endophenotypes. Yet, a multi-systemic analysis revealed the pleiotropic effects of Tanc2 outside the brain, such as growth failure and hepatocellular damage. This was associated with aberrant liver function including altered hepatocellular metabolism. Integrative analysis indicates that these disrupted Tanc2 systemic effects relate to interaction with Hippo developmental signalling pathway proteins and will increase the risk for comorbid somatic disease. This highlights how NDD gene pleiotropy can augment medical comorbidity susceptibility, underscoring the benefit of holistic NDD patient diagnosis and treatment for which large-scale preclinical functional genomics can provide complementary pleiotropic gene function information.
Keywords: Mouse models; Neurodevelopmental disorder; Somatic comorbidity; TANC2.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
References
-
- American Psychiatric Association (2013). Diagnostic and Statistical Manual of Mental Disorders, 5th edn. Arlington, VA: American Psychiatric Association.
-
- Bertomeu, T., Coulombe-Huntington, J., Chatr-Aryamontri, A., Bourdages, K. G., Coyaud, E., Raught, B., Xia, Y. and Tyers, M. (2018). A high-resolution genome-wide CRISPR/Cas9 viability screen reveals structural features and contextual diversity of the human cell-essential Proteome. Mol. Cell. Biol. 38, e00302-17. 10.1128/MCB.00302-17 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
