

Mucosal Vaccination with Cyclic Dinucleotide Adjuvants Induces Effective T Cell Homing and IL-17-Dependent Protection against Mycobacterium tuberculosis Infection
- PMID: 34965963
- PMCID: PMC8755605
- DOI: 10.4049/jimmunol.2100029
Mucosal Vaccination with Cyclic Dinucleotide Adjuvants Induces Effective T Cell Homing and IL-17-Dependent Protection against Mycobacterium tuberculosis Infection
Abstract
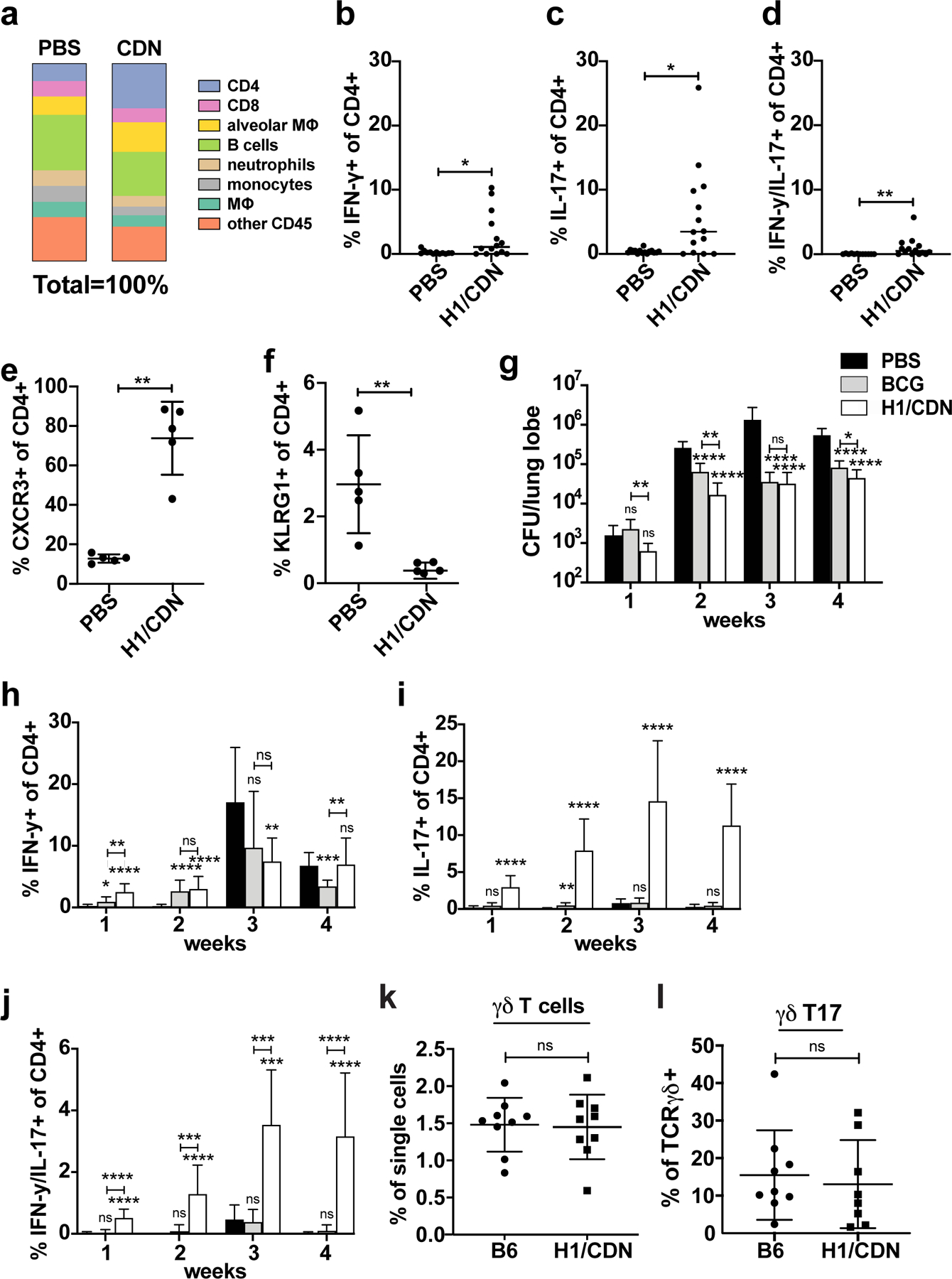
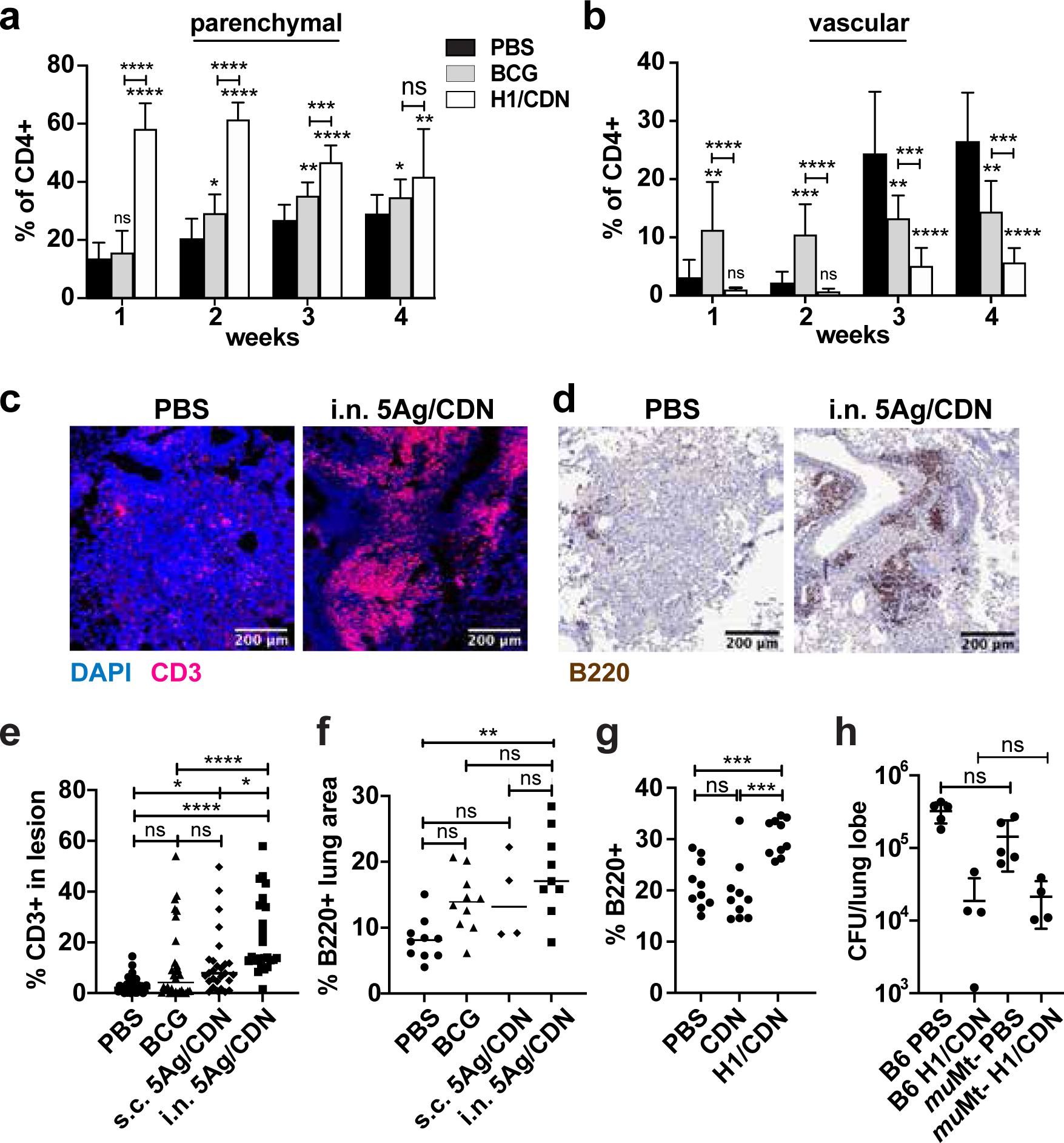
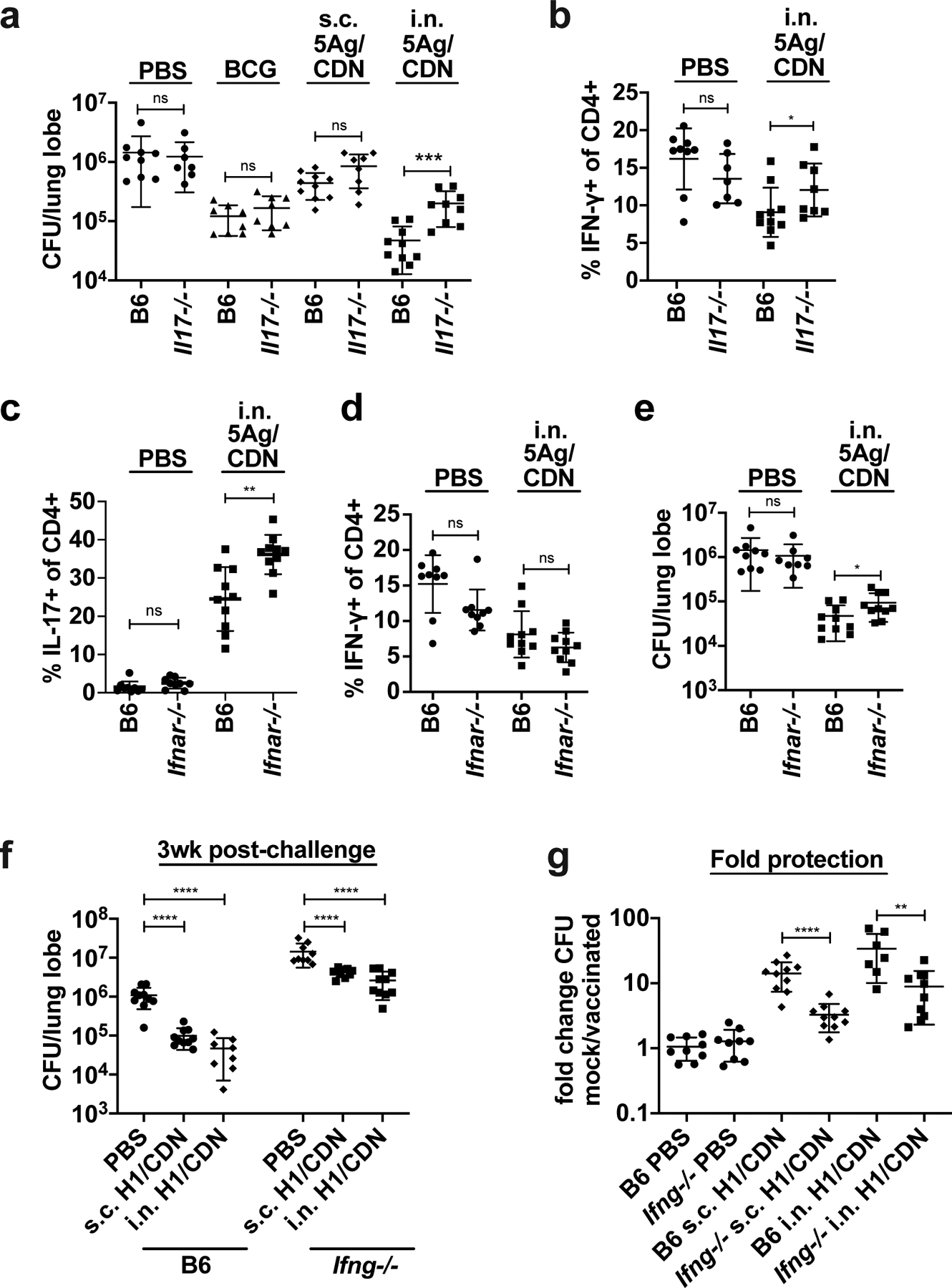
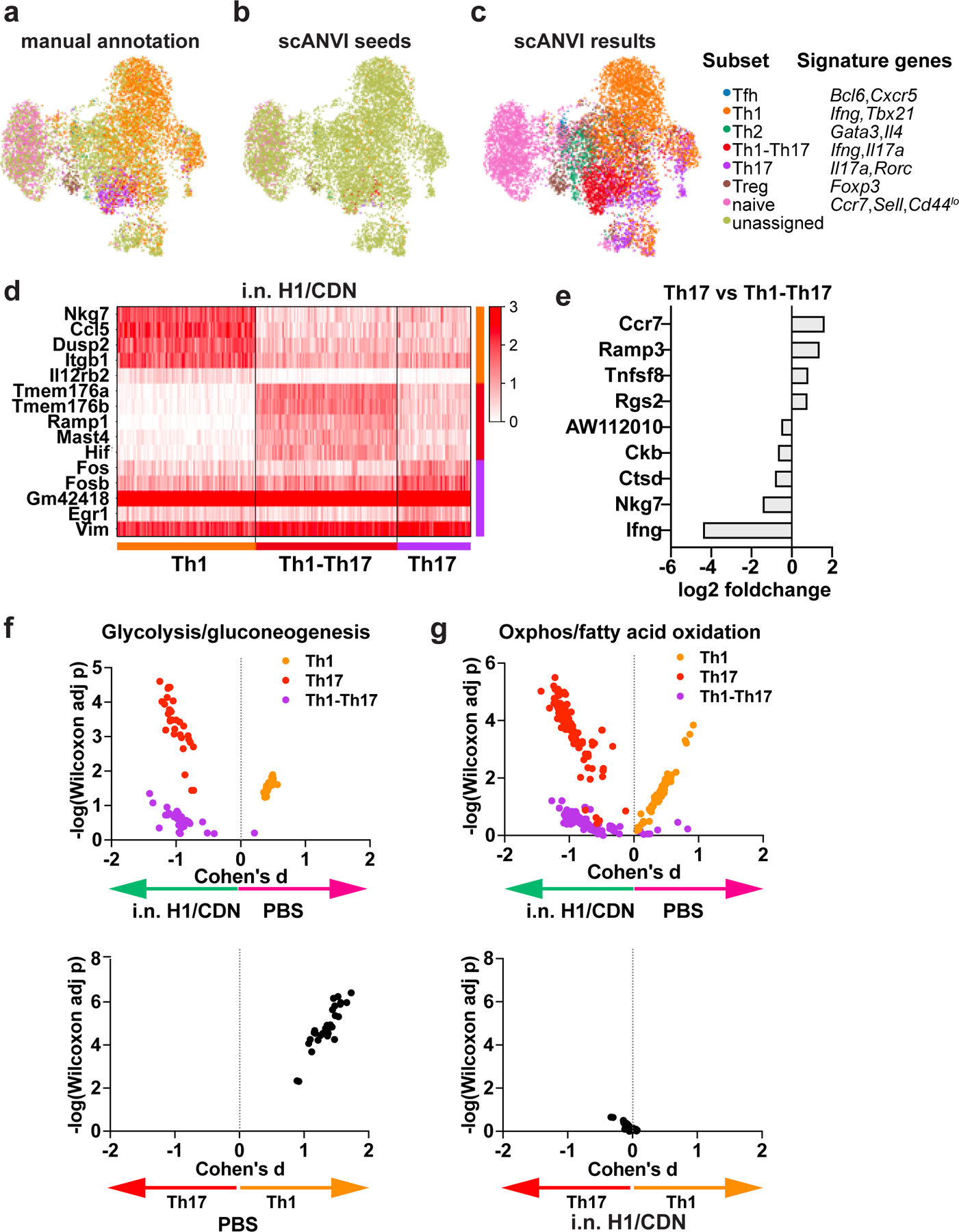
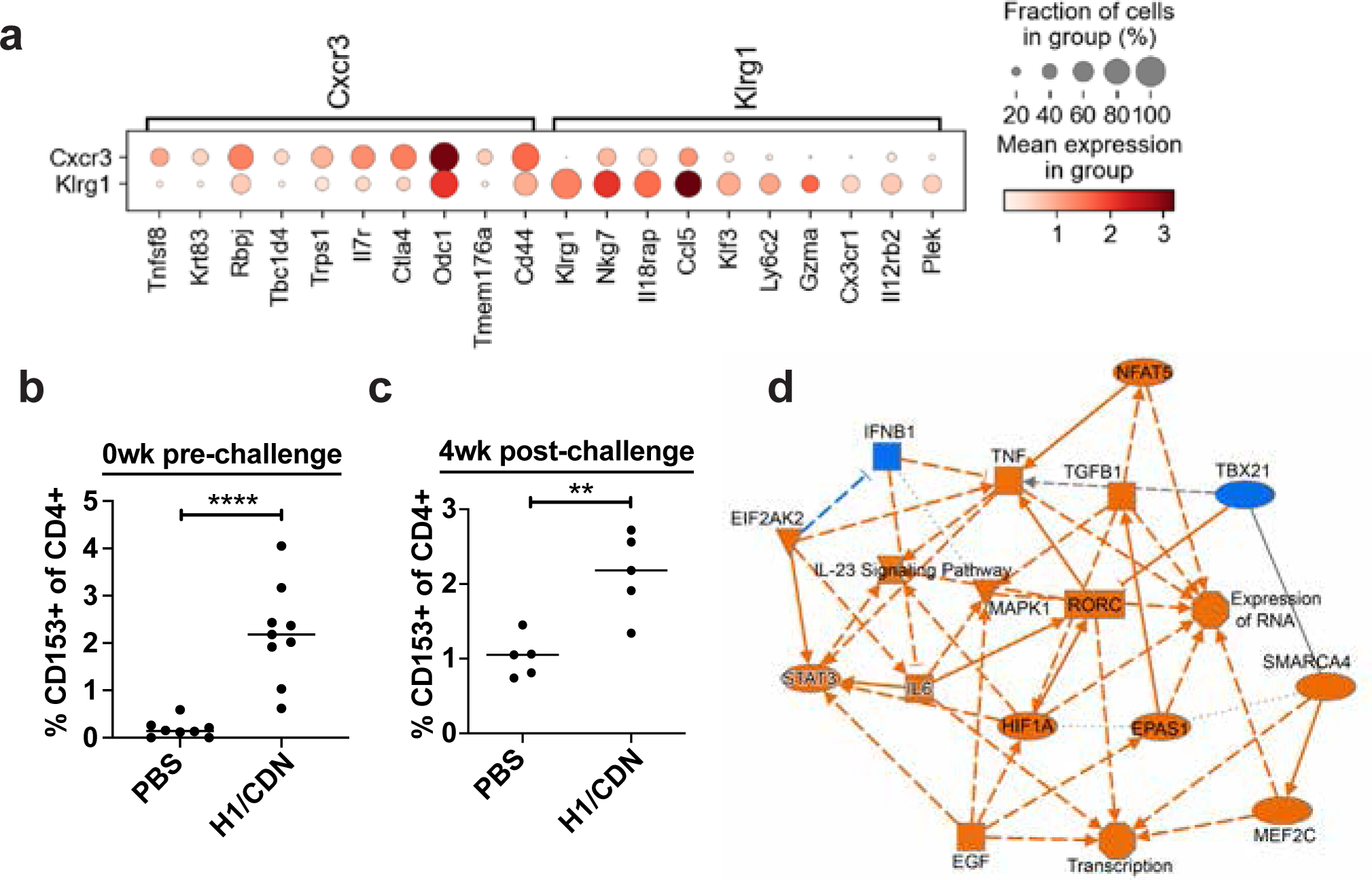
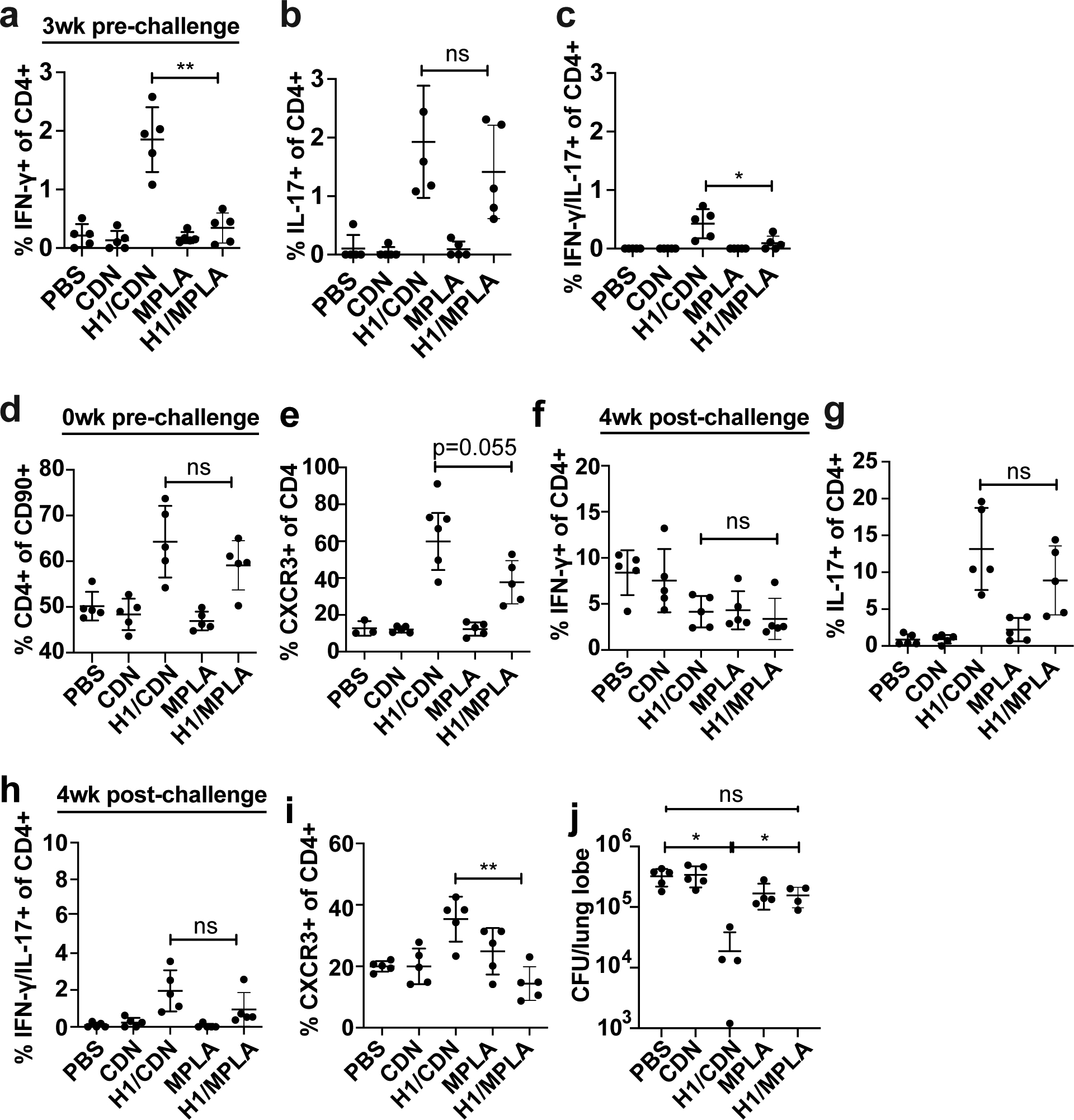
Tuberculosis consistently causes more deaths worldwide annually than any other single pathogen, making new effective vaccines an urgent priority for global public health. Among potential adjuvants, STING-activating cyclic dinucleotides (CDNs) uniquely stimulate a cytosolic sensing pathway activated only by pathogens. Recently, we demonstrated that a CDN-adjuvanted protein subunit vaccine robustly protects against tuberculosis infection in mice. In this study, we delineate the mechanistic basis underlying the efficacy of CDN vaccines for tuberculosis. CDN vaccines elicit CD4 T cells that home to lung parenchyma and penetrate into macrophage lesions in the lung. Although CDNs, like other mucosal vaccines, generate B cell-containing lymphoid structures in the lungs, protection is independent of B cells. Mucosal vaccination with a CDN vaccine induces Th1, Th17, and Th1-Th17 cells, and protection is dependent upon both IL-17 and IFN-γ. Single-cell RNA sequencing experiments reveal that vaccination enhances a metabolic state in Th17 cells reflective of activated effector function and implicate expression of Tnfsf8 (CD153) in vaccine-induced protection. Finally, we demonstrate that simply eliciting Th17 cells via mucosal vaccination with any adjuvant is not sufficient for protection. A vaccine adjuvanted with deacylated monophosphoryl lipid A (MPLA) failed to protect against tuberculosis infection when delivered mucosally, despite eliciting Th17 cells, highlighting the unique promise of CDNs as adjuvants for tuberculosis vaccines.
Copyright © 2022 by The American Association of Immunologists, Inc.
Figures
Similar articles
-
STING-Activating Adjuvants Elicit a Th17 Immune Response and Protect against Mycobacterium tuberculosis Infection.Cell Rep. 2018 May 1;23(5):1435-1447. doi: 10.1016/j.celrep.2018.04.003. Cell Rep. 2018. PMID: 29719256 Free PMC article.
-
Combinational PRR Agonists in Liposomal Adjuvant Enhances Immunogenicity and Protective Efficacy in a Tuberculosis Subunit Vaccine.Front Immunol. 2020 Sep 30;11:575504. doi: 10.3389/fimmu.2020.575504. eCollection 2020. Front Immunol. 2020. PMID: 33117374 Free PMC article.
-
Enhanced effect of BCG vaccine against pulmonary Mycobacterium tuberculosis infection in mice with lung Th17 response to mycobacterial heparin-binding hemagglutinin adhesin antigen.Microbiol Immunol. 2015 Dec;59(12):735-43. doi: 10.1111/1348-0421.12340. Microbiol Immunol. 2015. PMID: 26577130
-
Immunization strategies against pulmonary tuberculosis: considerations of T cell geography.Adv Exp Med Biol. 2013;783:267-78. doi: 10.1007/978-1-4614-6111-1_14. Adv Exp Med Biol. 2013. PMID: 23468114 Review.
-
[Novel vaccines against M. tuberculosis].Kekkaku. 2006 Dec;81(12):745-51. Kekkaku. 2006. PMID: 17240920 Review. Japanese.
Cited by
-
STING Targeting in Lung Diseases.Cells. 2022 Nov 3;11(21):3483. doi: 10.3390/cells11213483. Cells. 2022. PMID: 36359882 Free PMC article. Review.
-
Mycobacterium tuberculosis suppresses protective Th17 responses during infection through multiple mechanisms.bioRxiv [Preprint]. 2025 May 13:2025.05.08.652811. doi: 10.1101/2025.05.08.652811. bioRxiv. 2025. PMID: 40463021 Free PMC article. Preprint.
-
Mucosal immunotherapy targeting APC in lung disease.J Inflamm (Lond). 2025 Apr 14;22(1):15. doi: 10.1186/s12950-025-00432-2. J Inflamm (Lond). 2025. PMID: 40229816 Free PMC article. Review.
-
Protection of infant mice against pertussis, tuberculosis and influenza by co-administration of nasal pertussis vaccine candidate BPZE1 and BCG.iScience. 2025 Jun 7;28(7):112839. doi: 10.1016/j.isci.2025.112839. eCollection 2025 Jul 18. iScience. 2025. PMID: 40612502 Free PMC article.
-
A protective, single-visit TB vaccination regimen by co-administration of a subunit vaccine with BCG.NPJ Vaccines. 2023 May 9;8(1):66. doi: 10.1038/s41541-023-00666-2. NPJ Vaccines. 2023. PMID: 37160970 Free PMC article.
References
-
- Geneva. 2018. Global tuberculosis report 2018,. World Health Organization.
-
- Colditz GA 1994. The efficacy of bcg vaccine in the prevention of tuberculosis. Pediatr. Infect. Dis. J 13: 761.
-
- Fine PE 1995. Variation in protection by BCG: implications of and for heterologous immunity. Lancet 346: 1339–1345. - PubMed
-
- Mangtani P, Abubakar I, Ariti C, Beynon R, Pimpin L, Fine PEM, Rodrigues LC, Smith PG, Lipman M, Whiting PF, and Sterne JA. 2014. Protection by BCG vaccine against tuberculosis: a systematic review of randomized controlled trials. Clin. Infect. Dis 58: 470–480. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
