

Completing the family of human Eps15 homology domains: Solution structure of the internal Eps15 homology domain of γ-synergin
- PMID: 34967068
- PMCID: PMC8927860
- DOI: 10.1002/pro.4269
Completing the family of human Eps15 homology domains: Solution structure of the internal Eps15 homology domain of γ-synergin
Abstract
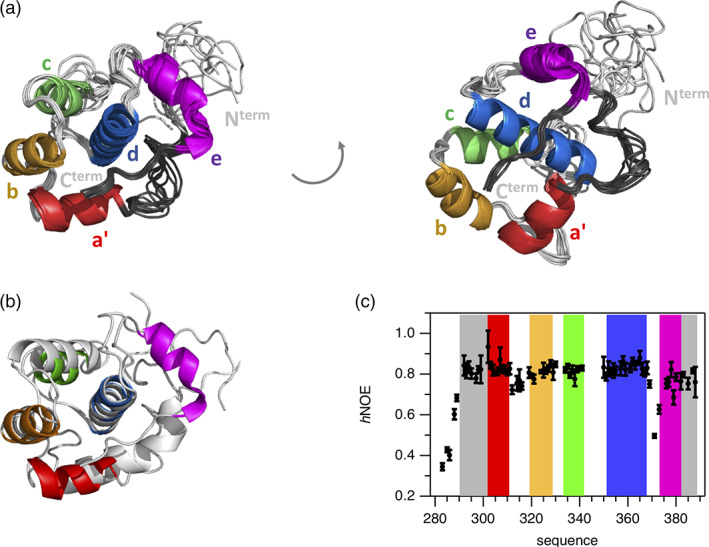
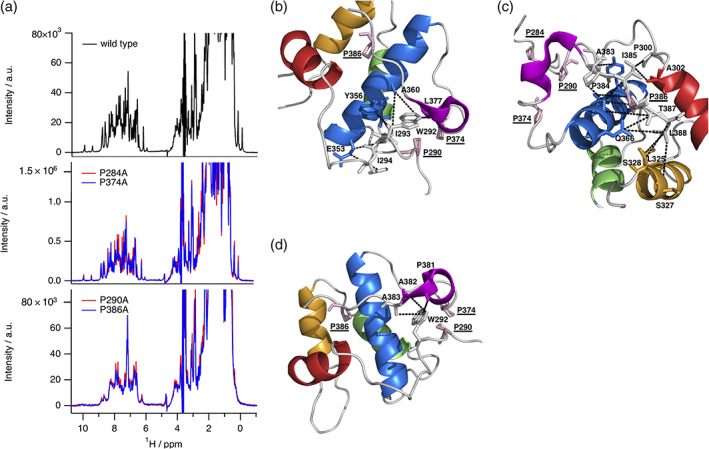
Eps15 homology (EH) domains are universal interaction domains to establish networks of protein-protein interactions in the cell. These networks mainly coordinate cellular functions including endocytosis, actin remodeling, and other intracellular signaling pathways. They are well characterized in structural terms, except for the internal EH domain from human γ-synergin (EHγ). Here, we complete the family of EH domain structures by determining the solution structure of the EHγ domain. The structural ensemble follows the canonical EH domain fold and the identified binding site is similar to other known EH domains. But EHγ differs significantly in the N- and C-terminal regions. The N-terminal α-helix is shortened compared to known homologues, while the C-terminal one is fully formed. A significant proportion of the remaining N- and C-terminal regions are well structured, a feature not seen in other EH domains. Single mutations in both the N-terminal and the C-terminal structured extensions lead to the loss of the distinct three-dimensional fold and turn EHγ into a molten globule like state. Therefore, we propose that the structural extensions in EHγ function as a clamp and are undoubtedly required to maintain its tertiary fold.
Keywords: EF hand; EH domain; Eps15 homology domain; NMR spectroscopy; molten globule; structure determination.
© 2021 European Molecular Biology Laboratory. Protein Science published by Wiley Periodicals LLC on behalf of The Protein Society.
Conflict of interest statement
Authors declare that they have no competing interests.
Figures



Similar articles
-
Mechanism for the selective interaction of C-terminal Eps15 homology domain proteins with specific Asn-Pro-Phe-containing partners.J Biol Chem. 2010 Mar 19;285(12):8687-94. doi: 10.1074/jbc.M109.045666. Epub 2010 Jan 27. J Biol Chem. 2010. PMID: 20106972 Free PMC article.
-
Solution structure of Eps15's third EH domain reveals coincident Phe-Trp and Asn-Pro-Phe binding sites.Biochemistry. 2000 Apr 18;39(15):4309-19. doi: 10.1021/bi9927383. Biochemistry. 2000. PMID: 10757979
-
EHD1 and Eps15 interact with phosphatidylinositols via their Eps15 homology domains.J Biol Chem. 2007 Jun 1;282(22):16612-22. doi: 10.1074/jbc.M609493200. Epub 2007 Apr 5. J Biol Chem. 2007. PMID: 17412695
-
The Eps15 homology (EH) domain.FEBS Lett. 2002 Feb 20;513(1):24-9. doi: 10.1016/s0014-5793(01)03241-0. FEBS Lett. 2002. PMID: 11911876 Review.
-
EH and UIM: endocytosis and more.Sci STKE. 2003 Dec 16;2003(213):re17. doi: 10.1126/stke.2132003re17. Sci STKE. 2003. PMID: 14679291 Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
