

Redirecting meiotic DNA break hotspot determinant proteins alters localized spatial control of DNA break formation and repair
- PMID: 34967417
- PMCID: PMC8789058
- DOI: 10.1093/nar/gkab1253
Redirecting meiotic DNA break hotspot determinant proteins alters localized spatial control of DNA break formation and repair
Abstract
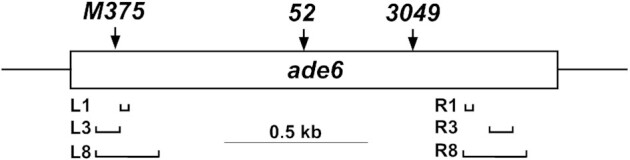
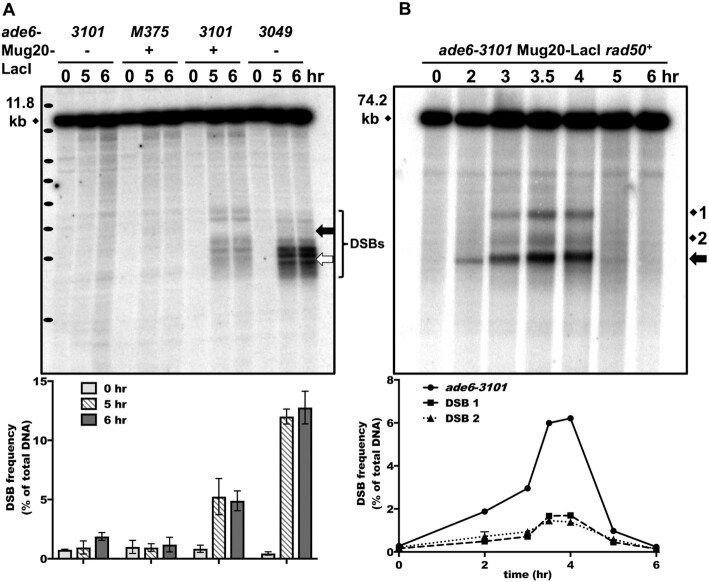
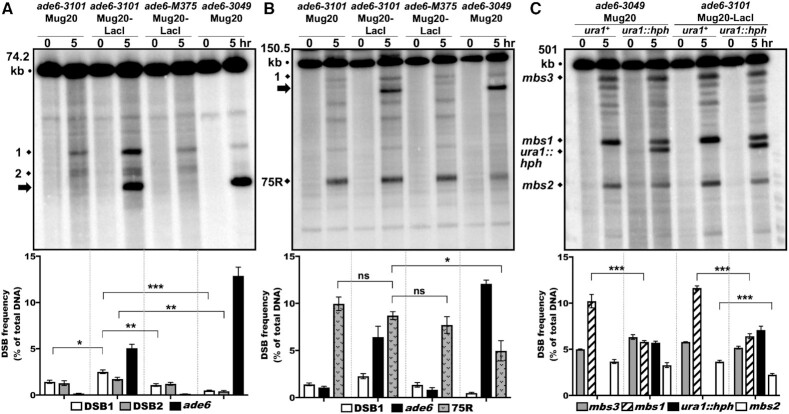
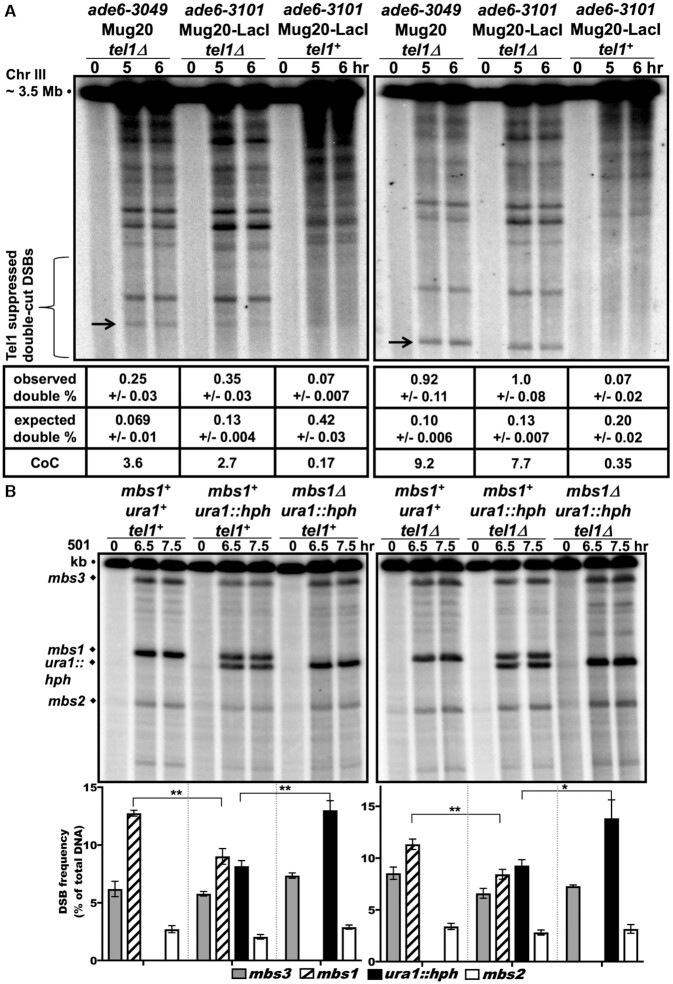
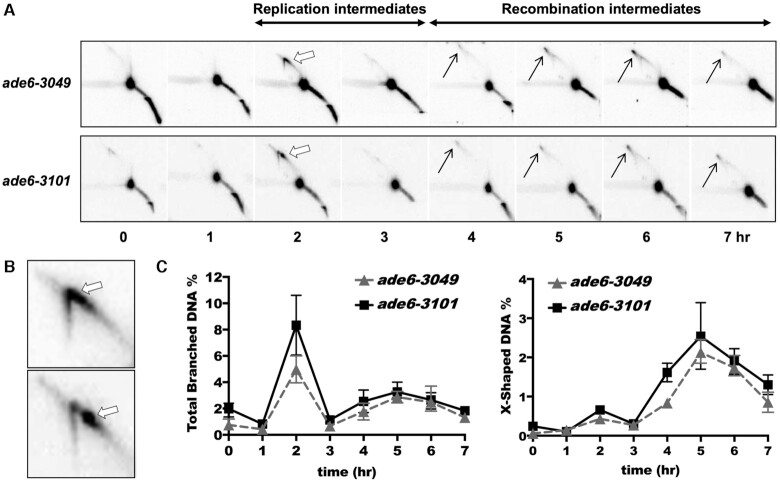
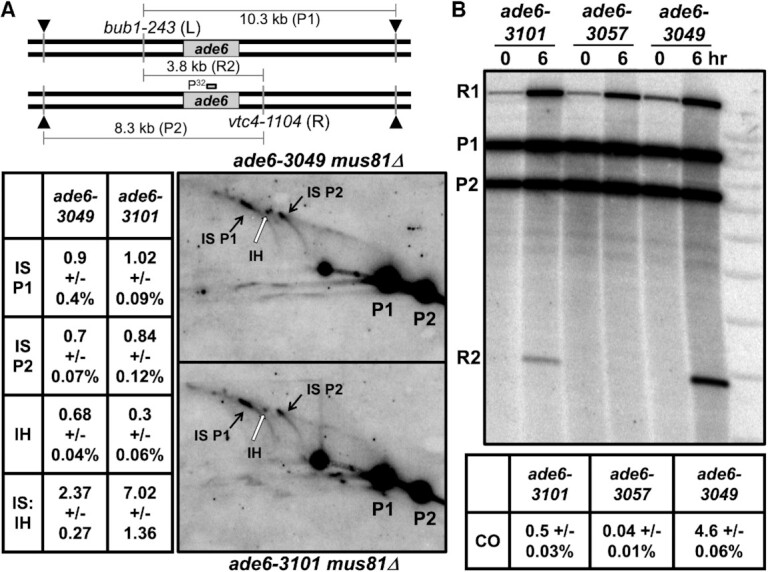
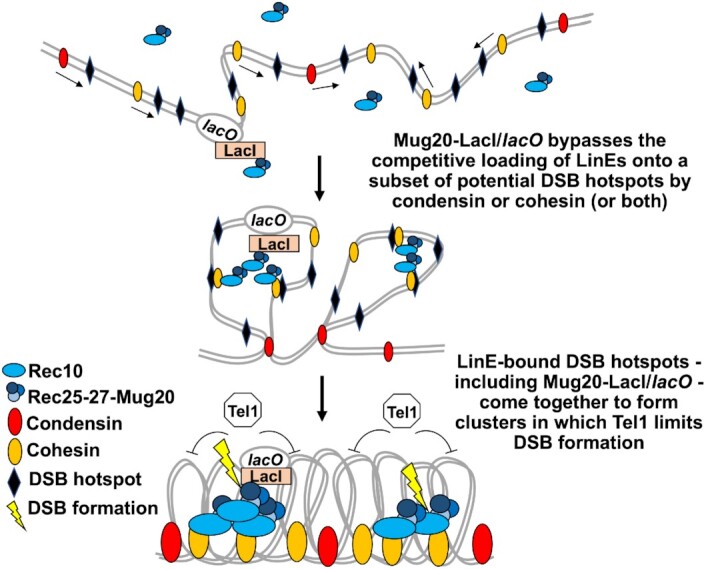
During meiosis, DNA double-strand breaks (DSBs) are formed at high frequency at special chromosomal sites, called DSB hotspots, to generate crossovers that aid proper chromosome segregation. Multiple chromosomal features affect hotspot formation. In the fission yeast S. pombe the linear element proteins Rec25, Rec27 and Mug20 are hotspot determinants - they bind hotspots with high specificity and are necessary for nearly all DSBs at hotspots. To assess whether they are also sufficient for hotspot determination, we localized each linear element protein to a novel chromosomal site (ade6 with lacO substitutions) by fusion to the Escherichia coli LacI repressor. The Mug20-LacI plus lacO combination, but not the two separate lac elements, produced a strong ade6 DSB hotspot, comparable to strong endogenous DSB hotspots. This hotspot had unexpectedly low ade6 recombinant frequency and negligible DSB hotspot competition, although like endogenous hotspots it manifested DSB interference. We infer that linear element proteins must be properly placed by endogenous functions to impose hotspot competition and proper partner choice for DSB repair. Our results support and expand our previously proposed DSB hotspot-clustering model for local control of meiotic recombination.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Zickler D., Kleckner N.. Meiotic chromosomes: integrating structure and function. Annu. Rev. Genet. 1999; 33:603–754. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
