

ABI1-based expression signature predicts breast cancer metastasis and survival
- PMID: 34967509
- PMCID: PMC9297774
- DOI: 10.1002/1878-0261.13175
ABI1-based expression signature predicts breast cancer metastasis and survival
Abstract
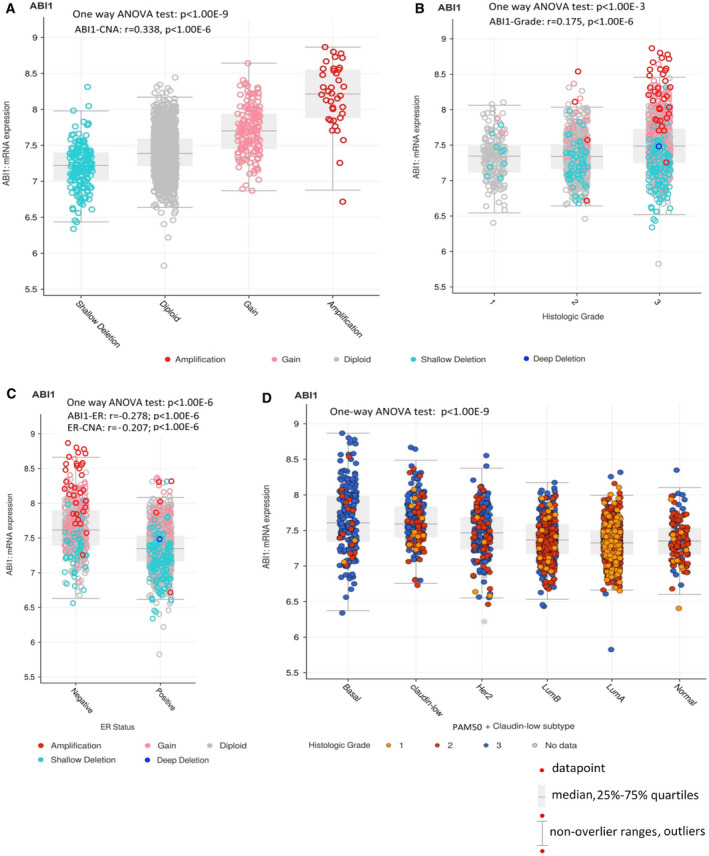
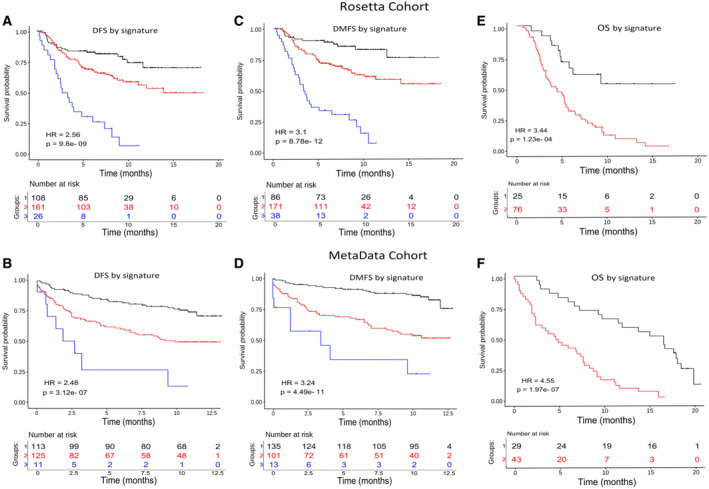
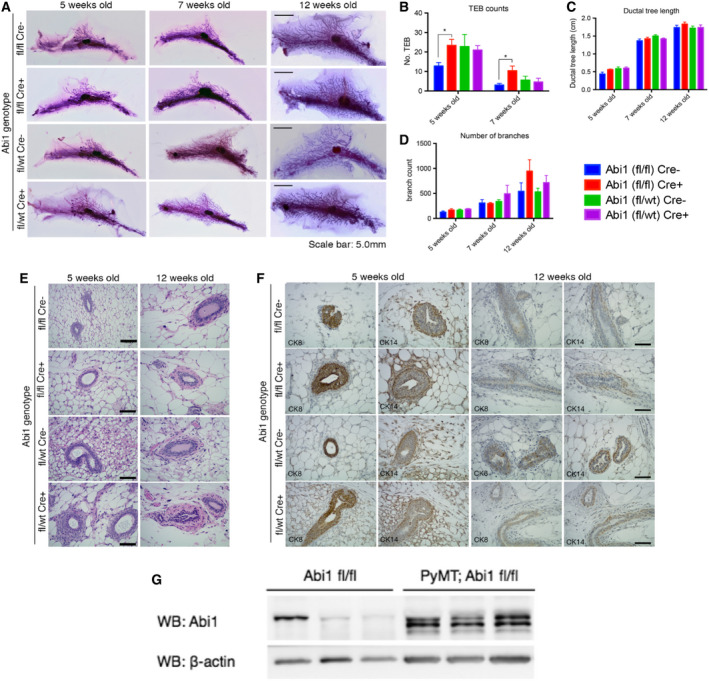
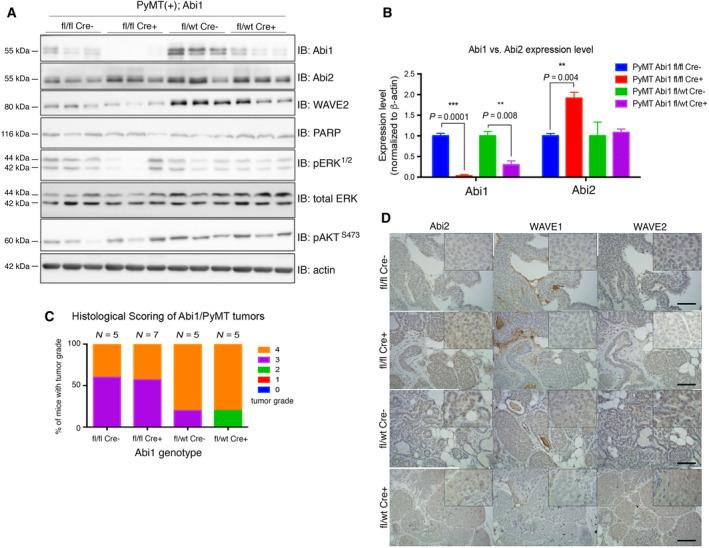
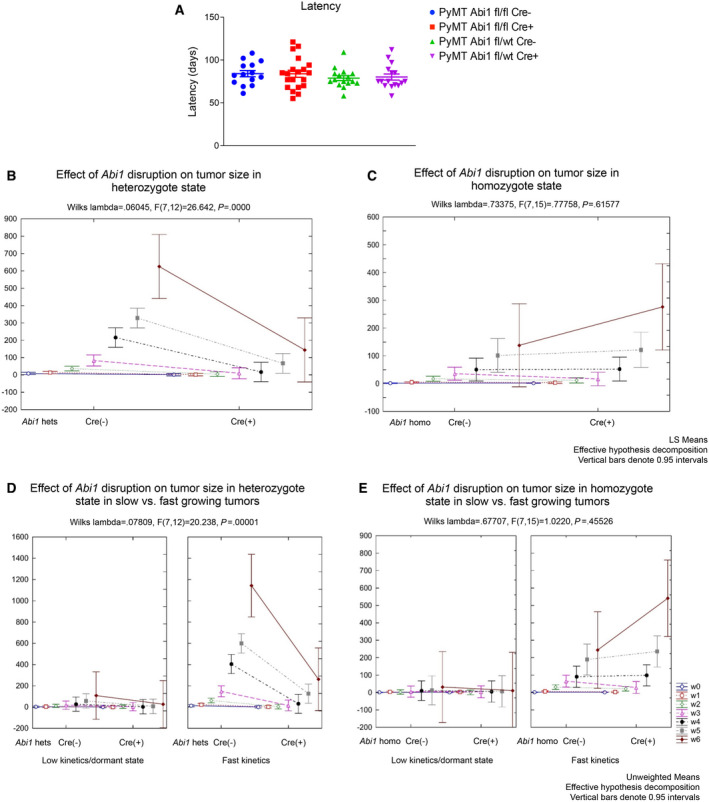
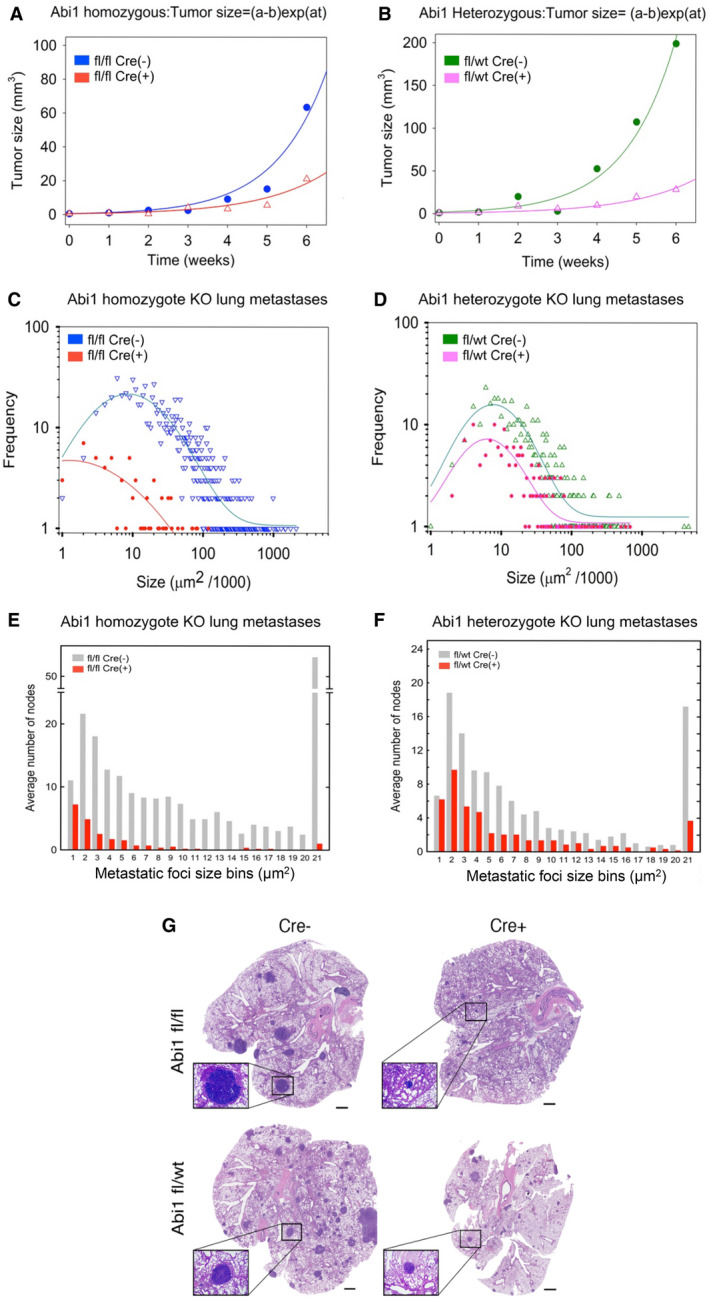
Despite the current standard of care, breast cancer remains one of the leading causes of mortality in women worldwide, thus emphasizing the need for better predictive and therapeutic targets. ABI1 is associated with poor survival and an aggressive breast cancer phenotype, although its role in tumorigenesis, metastasis, and the disease outcome remains to be elucidated. Here, we define the ABI1-based seven-gene prognostic signature that predicts survival of metastatic breast cancer patients; ABI1 is an essential component of the signature. Genetic disruption of Abi1 in primary breast cancer tumors of PyMT mice led to significant reduction of the number and size of lung metastases in a gene dose-dependent manner. The disruption of Abi1 resulted in deregulation of the WAVE complex at the mRNA and protein levels in mouse tumors. In conclusion, ABI1 is a prognostic metastatic biomarker in breast cancer. We demonstrate, for the first time, that lung metastasis is associated with an Abi1 gene dose and specific gene expression aberrations in primary breast cancer tumors. These results indicate that targeting ABI1 may provide a therapeutic advantage in breast cancer patients.
Keywords: Abi1; WAVE; breast cancer; metastasis; preclinical mouse; survival signature.
© 2021 The Authors. Molecular Oncology published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
