

Sod1 integrates oxygen availability to redox regulate NADPH production and the thiol redoxome
- PMID: 34969852
- PMCID: PMC8740578
- DOI: 10.1073/pnas.2023328119
Sod1 integrates oxygen availability to redox regulate NADPH production and the thiol redoxome
Abstract
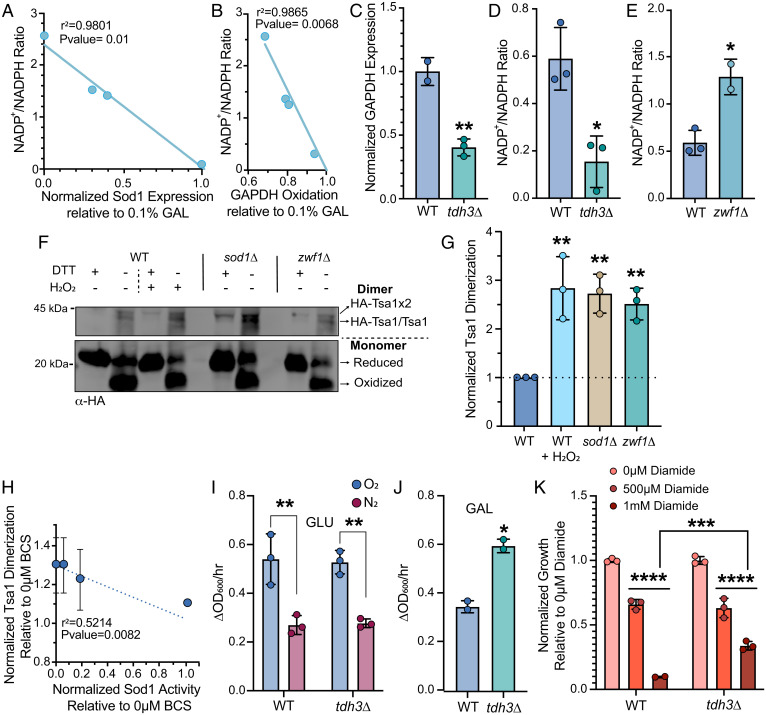
Cu/Zn superoxide dismutase (Sod1) is a highly conserved and abundant antioxidant enzyme that detoxifies superoxide (O2•-) by catalyzing its conversion to dioxygen (O2) and hydrogen peroxide (H2O2). Using Saccharomyces cerevisiae and mammalian cells, we discovered that a major aspect of the antioxidant function of Sod1 is to integrate O2 availability to promote NADPH production. The mechanism involves Sod1-derived H2O2 oxidatively inactivating the glycolytic enzyme, GAPDH, which in turn reroutes carbohydrate flux to the oxidative phase of the pentose phosphate pathway (oxPPP) to generate NADPH. The aerobic oxidation of GAPDH is dependent on and rate-limited by Sod1. Thus, Sod1 senses O2 via O2•- to balance glycolytic and oxPPP flux, through control of GAPDH activity, for adaptation to life in air. Importantly, this mechanism for Sod1 antioxidant activity requires the bulk of cellular Sod1, unlike for its role in protection against O2•- toxicity, which only requires <1% of total Sod1. Using mass spectrometry, we identified proteome-wide targets of Sod1-dependent redox signaling, including numerous metabolic enzymes. Altogether, Sod1-derived H2O2 is important for antioxidant defense and a master regulator of metabolism and the thiol redoxome.
Keywords: glycolysis; oxygen sensing; pentose phosphate pathway; redox signaling; superoxide dismutase.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures




References
-
- Flint D. H., Tuminello J. F., Emptage M. H., The inactivation of Fe-S cluster containing hydro-lyases by superoxide. J. Biol. Chem. 268, 22369–22376 (1993). - PubMed
-
- Liochev S. I., Fridovich I., Superoxide and iron: Partners in crime. IUBMB Life 48, 157–161 (1999). - PubMed
-
- Imlay J. A., Pathways of oxidative damage. Annu. Rev. Microbiol. 57, 395–418 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
