

Biophysical and biochemical properties of PHGDH revealed by studies on PHGDH inhibitors
- PMID: 34971423
- PMCID: PMC11073335
- DOI: 10.1007/s00018-021-04022-2
Biophysical and biochemical properties of PHGDH revealed by studies on PHGDH inhibitors
Abstract
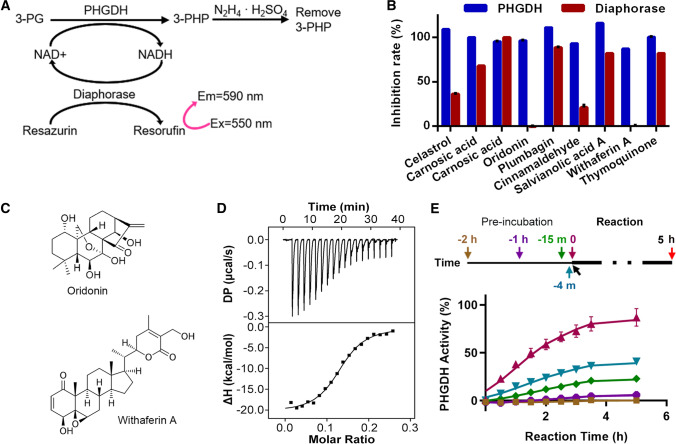
The rate-limiting serine biogenesis enzyme PHGDH is overexpressed in cancers. Both serine withdrawal and genetic/pharmacological inhibition of PHGDH have demonstrated promising tumor-suppressing activities. However, the enzyme properties of PHGDH are not well understood and the discovery of PHGDH inhibitors is still in its infancy. Here, oridonin was identified from a natural product library as a new PHGDH inhibitor. The crystal structure of PHGDH in complex with oridonin revealed a new allosteric site. The binding of oridonin to this site reduced the activity of the enzyme by relocating R54, a residue involved in substrate binding. Mutagenesis studies showed that PHGDH activity was very sensitive to cysteine mutations, especially those in the substrate binding domain. Conjugation of oridonin and other reported covalent PHGDH inhibitors to these sites will therefore inhibit PHGDH. In addition to being inhibited enzymatically, PHGDH can also be inhibited by protein aggregation and proteasome-mediated degradation. Several tested PHGDH cancer mutants showed altered enzymatic activity, which can be explained by protein structure and stability. Overall, the above studies present new biophysical and biochemical insights into PHGDH and may facilitate the future design of PHGDH inhibitors.
Keywords: Allosteric inhibition; Cancer mutation; Compound screening; Nucleotide-binding domain; Protein degradation.
© 2021. The Author(s), under exclusive licence to Springer Nature Switzerland AG.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures





Similar articles
-
Inhibition of 3-phosphoglycerate dehydrogenase (PHGDH) by indole amides abrogates de novo serine synthesis in cancer cells.Bioorg Med Chem Lett. 2019 Sep 1;29(17):2503-2510. doi: 10.1016/j.bmcl.2019.07.011. Epub 2019 Jul 6. Bioorg Med Chem Lett. 2019. PMID: 31327531 Free PMC article.
-
Rational Design of Selective Allosteric Inhibitors of PHGDH and Serine Synthesis with Anti-tumor Activity.Cell Chem Biol. 2017 Jan 19;24(1):55-65. doi: 10.1016/j.chembiol.2016.11.013. Epub 2016 Dec 29. Cell Chem Biol. 2017. PMID: 28042046 Free PMC article.
-
A retrospective overview of PHGDH and its inhibitors for regulating cancer metabolism.Eur J Med Chem. 2021 May 5;217:113379. doi: 10.1016/j.ejmech.2021.113379. Epub 2021 Mar 16. Eur J Med Chem. 2021. PMID: 33756126 Review.
-
Unravelling the Allosteric Targeting of PHGDH at the ACT-Binding Domain with a Photoactivatable Diazirine Probe and Mass Spectrometry Experiments.Molecules. 2021 Jan 18;26(2):477. doi: 10.3390/molecules26020477. Molecules. 2021. PMID: 33477510 Free PMC article.
-
Phosphoglycerate dehydrogenase (PHGDH) inhibitors: a comprehensive review 2015-2020.Expert Opin Ther Pat. 2021 Jul;31(7):597-608. doi: 10.1080/13543776.2021.1890028. Epub 2021 Mar 29. Expert Opin Ther Pat. 2021. PMID: 33571419 Review.
Cited by
-
Cycling back to folate metabolism in cancer.Nat Cancer. 2024 May;5(5):701-715. doi: 10.1038/s43018-024-00739-8. Epub 2024 May 2. Nat Cancer. 2024. PMID: 38698089 Free PMC article. Review.
-
PRMT1 Sustains De Novo Fatty Acid Synthesis by Methylating PHGDH to Drive Chemoresistance in Triple-Negative Breast Cancer.Cancer Res. 2024 Apr 1;84(7):1065-1083. doi: 10.1158/0008-5472.CAN-23-2266. Cancer Res. 2024. PMID: 38383964 Free PMC article.
-
The role of serine metabolism in lung cancer: From oncogenesis to tumor treatment.Front Genet. 2023 Jan 9;13:1084609. doi: 10.3389/fgene.2022.1084609. eCollection 2022. Front Genet. 2023. PMID: 36699468 Free PMC article. Review.
-
Metabolomic analysis of vascular cognitive impairment due to hepatocellular carcinoma.Front Neurol. 2023 Mar 16;13:1109019. doi: 10.3389/fneur.2022.1109019. eCollection 2022. Front Neurol. 2023. PMID: 37008043 Free PMC article.
-
Molecular glue triggers degradation of PHGDH by enhancing the interaction between DDB1 and PHGDH.Acta Pharm Sin B. 2024 Sep;14(9):4001-4013. doi: 10.1016/j.apsb.2024.06.001. Epub 2024 Jun 6. Acta Pharm Sin B. 2024. PMID: 39309493 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
