

Enzymatic and structural characterization of HAD5, an essential phosphomannomutase of malaria-causing parasites
- PMID: 34973333
- PMCID: PMC8808168
- DOI: 10.1016/j.jbc.2021.101550
Enzymatic and structural characterization of HAD5, an essential phosphomannomutase of malaria-causing parasites
Abstract
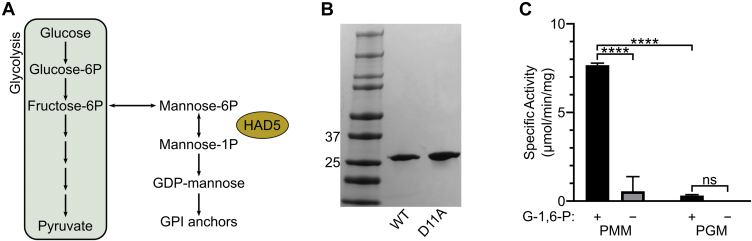
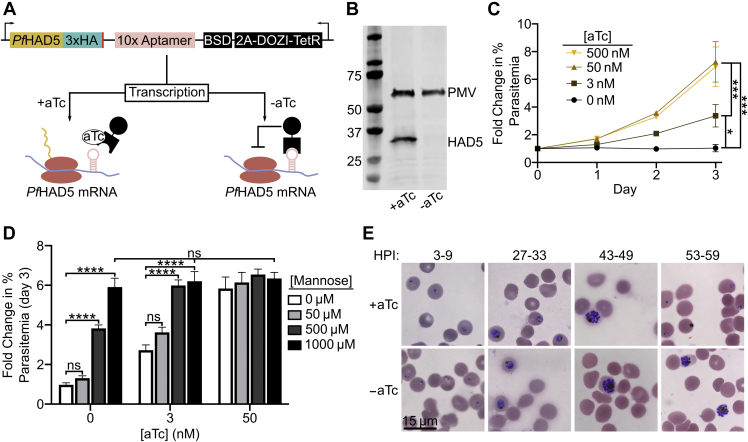
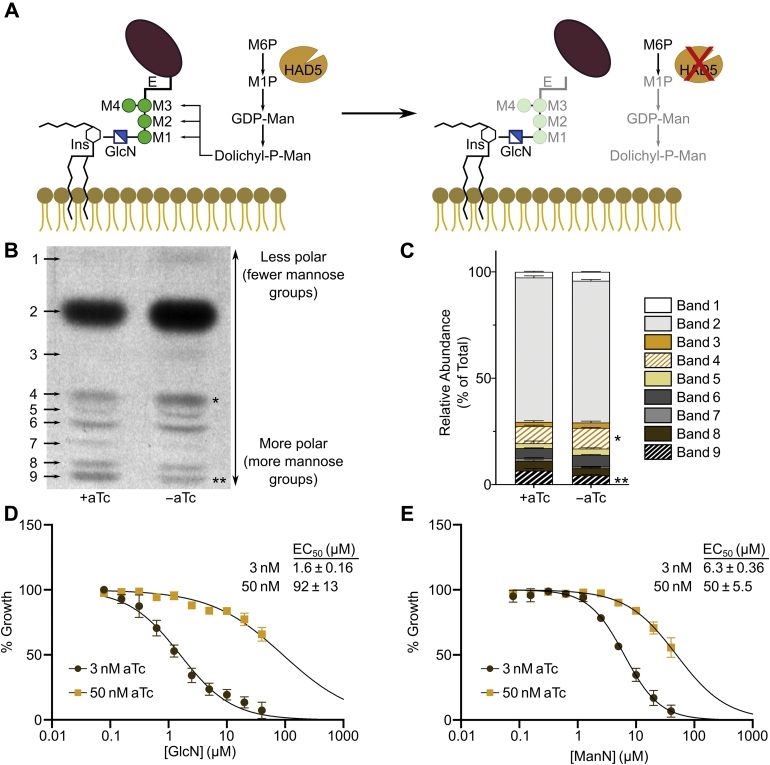
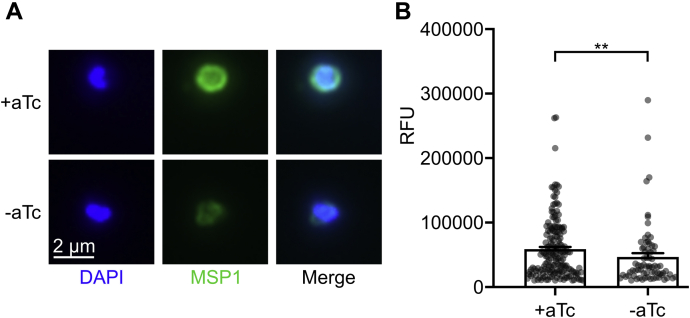
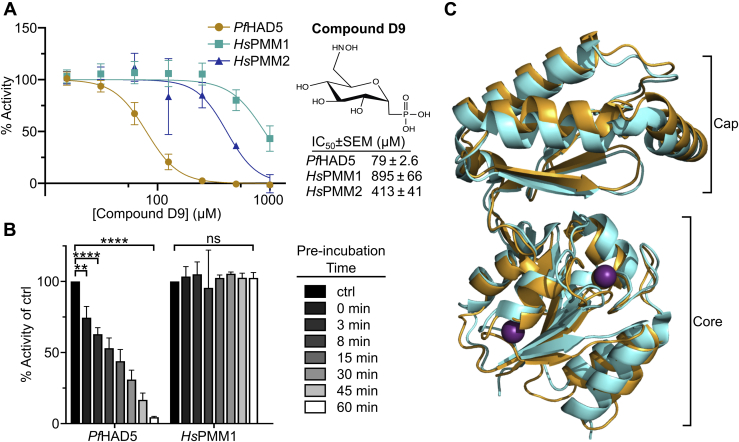
The malaria-causing parasite Plasmodium falciparum is responsible for over 200 million infections and 400,000 deaths per year. At multiple stages during its complex life cycle, P. falciparum expresses several essential proteins tethered to its surface by glycosylphosphatidylinositol (GPI) anchors, which are critical for biological processes such as parasite egress and reinvasion of host red blood cells. Targeting this pathway therapeutically has the potential to broadly impact parasite development across several life stages. Here, we characterize an upstream component of parasite GPI anchor biosynthesis, the putative phosphomannomutase (PMM) (EC 5.4.2.8), HAD5 (PF3D7_1017400). We confirmed the PMM and phosphoglucomutase activities of purified recombinant HAD5 by developing novel linked enzyme biochemical assays. By regulating the expression of HAD5 in transgenic parasites with a TetR-DOZI-inducible knockdown system, we demonstrated that HAD5 is required for malaria parasite egress and erythrocyte reinvasion, and we assessed the role of HAD5 in GPI anchor synthesis by autoradiography of radiolabeled glucosamine and thin layer chromatography. Finally, we determined the three-dimensional X-ray crystal structure of HAD5 and identified a substrate analog that specifically inhibits HAD5 compared to orthologous human PMMs in a time-dependent manner. These findings demonstrate that the GPI anchor biosynthesis pathway is exceptionally sensitive to inhibition in parasites and that HAD5 has potential as a specific, multistage antimalarial target.
Keywords: carbohydrate metabolism; crystal structure; drug development; glycosylphosphatidylinositol (GPI) anchor; haloacid dehalogenase (HAD); malaria; parasite; phosphomannomutase.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest A. R. O. J. reports financial support was provided by National Institutes of Health and Burroughs Wellcome Fund. D. L. J. reports financial support was provided by Natural Sciences and Engineering Research Council of Canada and Canadian Institutes of Health Research. A. R. O. J. reports a relationship with Pluton Biosciences that includes board membership and with American Society for Microbiology that includes consulting or advisory. A. R. O. J. has patent issued and pending patents on antimalarials pending to none.
Figures
References
-
- Yeung S. Malaria - update on antimalarial resistance and treatment approaches. Pediatr. Infect. Dis. J. 2018;37:367–369. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
