

The evolutionary history of the polyQ tract in huntingtin sheds light on its functional pro-neural activities
- PMID: 34974533
- PMCID: PMC8817008
- DOI: 10.1038/s41418-021-00914-9
The evolutionary history of the polyQ tract in huntingtin sheds light on its functional pro-neural activities
Abstract
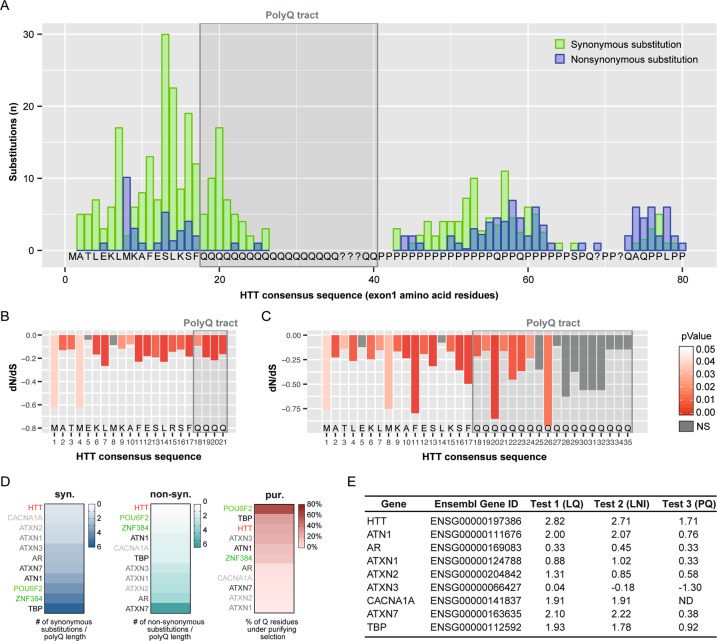
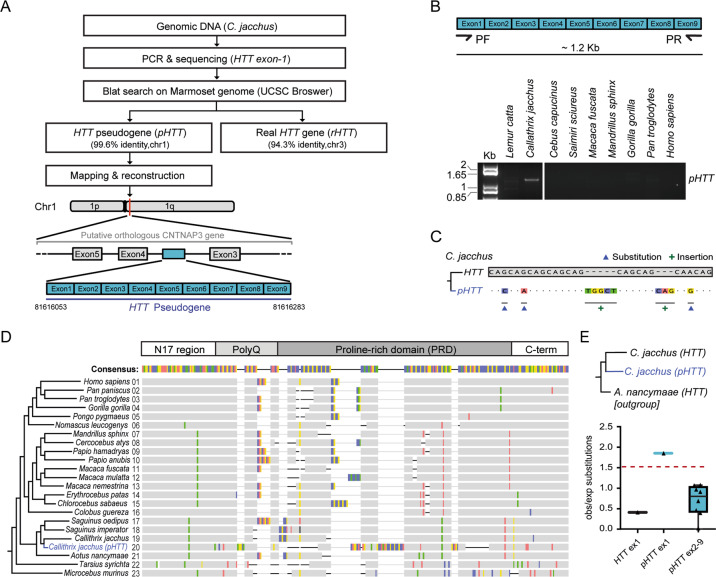
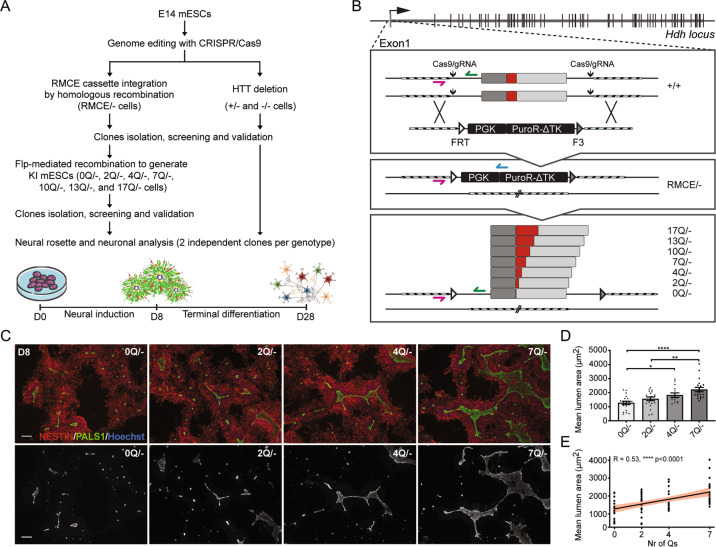
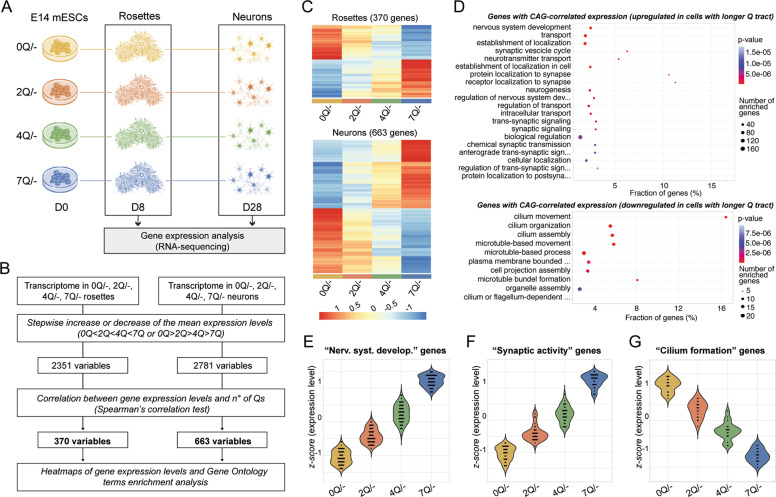
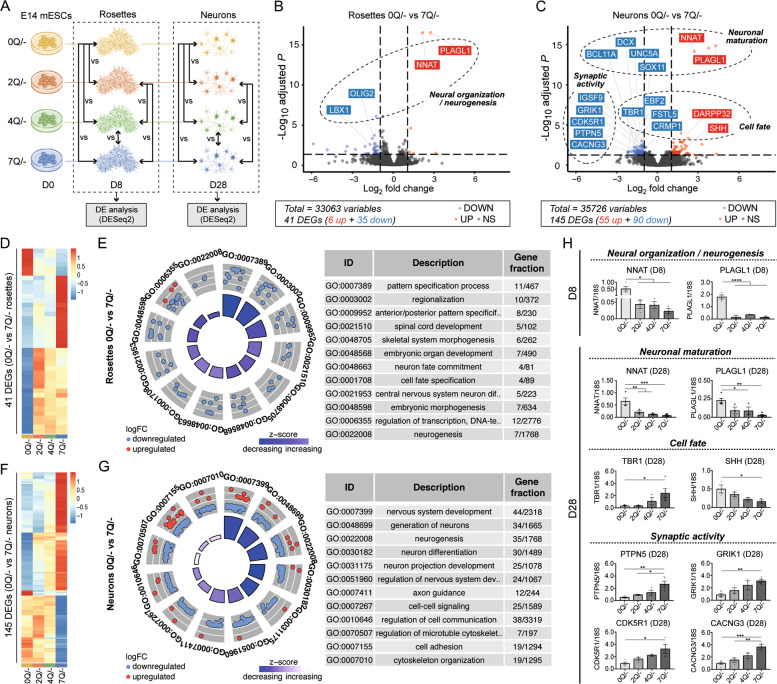
Huntington's disease is caused by a pathologically long (>35) CAG repeat located in the first exon of the Huntingtin gene (HTT). While pathologically expanded CAG repeats are the focus of extensive investigations, non-pathogenic CAG tracts in protein-coding genes are less well characterized. Here, we investigated the function and evolution of the physiological CAG tract in the HTT gene. We show that the poly-glutamine (polyQ) tract encoded by CAGs in the huntingtin protein (HTT) is under purifying selection and subjected to stronger selective pressures than CAG-encoded polyQ tracts in other proteins. For natural selection to operate, the polyQ must perform a function. By combining genome-edited mouse embryonic stem cells and cell assays, we show that small variations in HTT polyQ lengths significantly correlate with cells' neurogenic potential and with changes in the gene transcription network governing neuronal function. We conclude that during evolution natural selection promotes the conservation and purity of the CAG-encoded polyQ tract and that small increases in its physiological length influence neural functions of HTT. We propose that these changes in HTT polyQ length contribute to evolutionary fitness including potentially to the development of a more complex nervous system.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Extreme conservation of the poly-glutamine tract in huntingtin is related to neurodevelopmental functions: the "better" may become the "enemy of the good" in the course of evolution.Cell Death Differ. 2022 Feb;29(2):266-268. doi: 10.1038/s41418-021-00927-4. Epub 2022 Jan 10. Cell Death Differ. 2022. PMID: 35013554 Free PMC article. No abstract available.
References
-
- O'Donovan, MC. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. The Huntington’s Disease Collaborative Research Group. Cell. 1993;72:971–83. - PubMed
-
- Davies SW, Turmaine M, Cozens BA, DiFiglia M, Sharp AH, Ross CA, et al. Formation of neuronal intranuclear inclusions underlies the neurological dysfunction in mice transgenic for the HD mutation. Cell. 1997;90:537–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
