

Multidimensional Dynamics of the Proteome in the Neurodegenerative and Aging Mammalian Brain
- PMID: 34979241
- PMCID: PMC8816717
- DOI: 10.1016/j.mcpro.2021.100192
Multidimensional Dynamics of the Proteome in the Neurodegenerative and Aging Mammalian Brain
Abstract
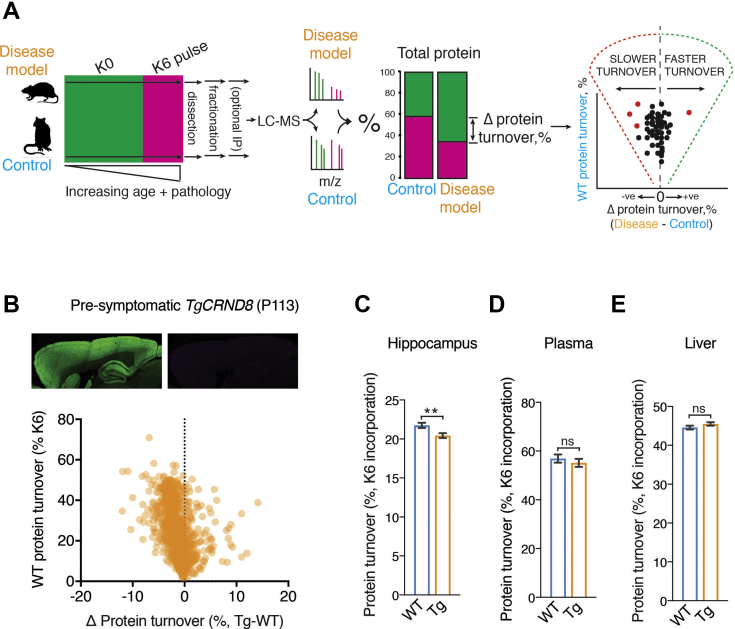
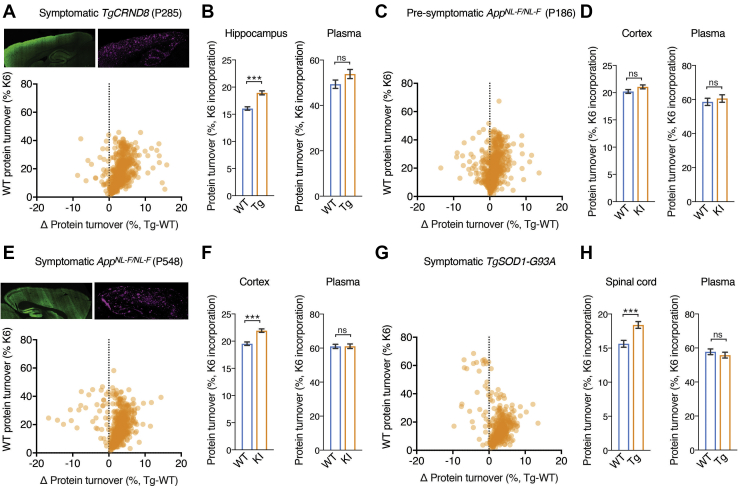
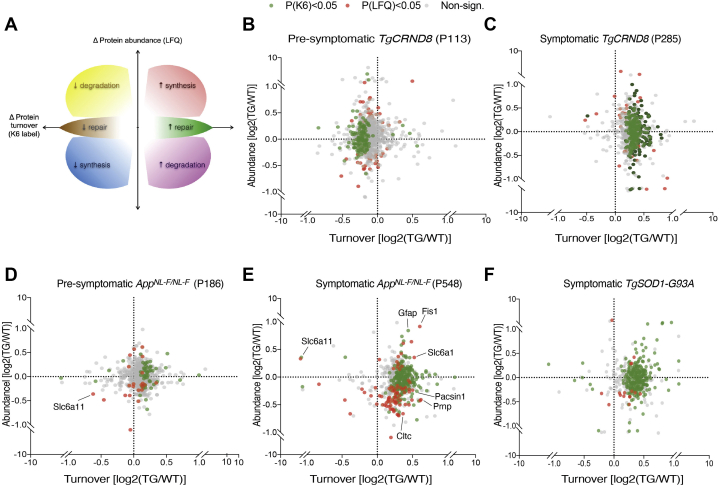
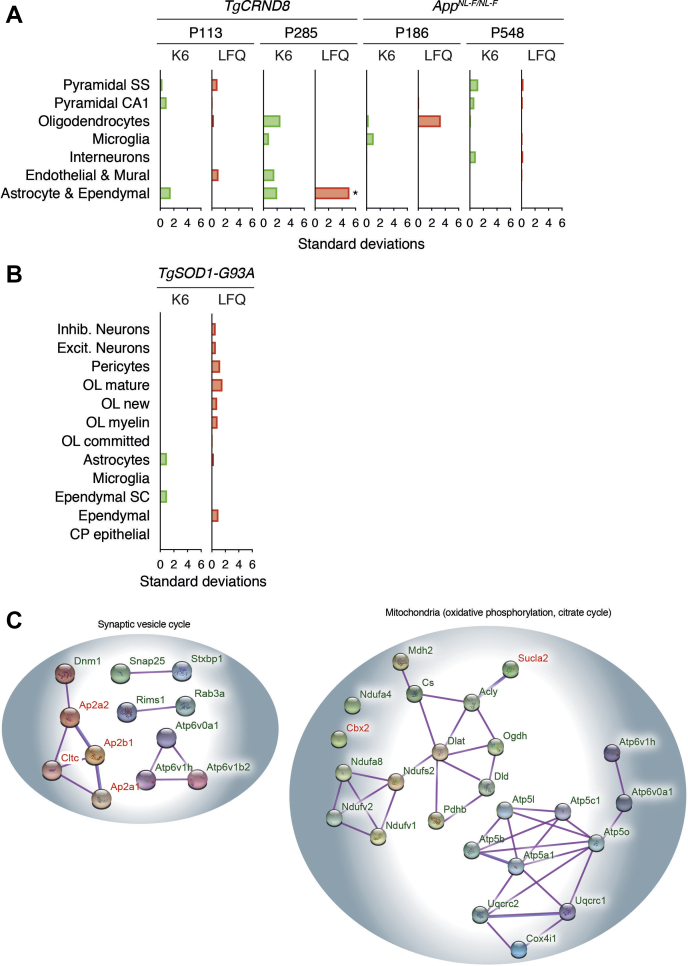
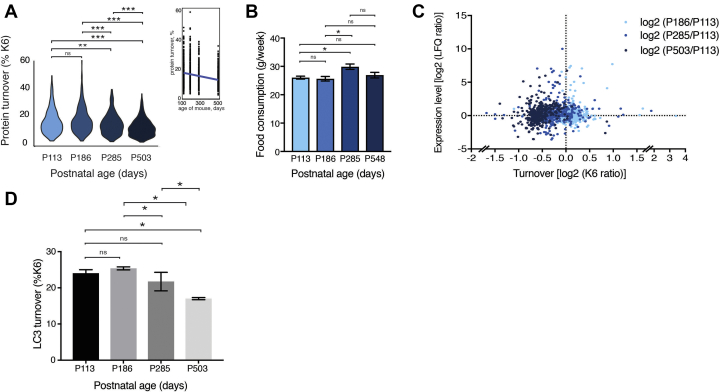
The amount of any given protein in the brain is determined by the rates of its synthesis and destruction, which are regulated by different cellular mechanisms. Here, we combine metabolic labeling in live mice with global proteomic profiling to simultaneously quantify both the flux and amount of proteins in mouse models of neurodegeneration. In multiple models, protein turnover increases were associated with increasing pathology. This method distinguishes changes in protein expression mediated by synthesis from those mediated by degradation. In the AppNL-F knockin mouse model of Alzheimer's disease, increased turnover resulted from imbalances in both synthesis and degradation, converging on proteins associated with synaptic vesicle recycling (Dnm1, Cltc, Rims1) and mitochondria (Fis1, Ndufv1). In contrast to disease models, aging in wild-type mice caused a widespread decrease in protein recycling associated with a decrease in autophagic flux. Overall, this simple multidimensional approach enables a comprehensive mapping of proteome dynamics and identifies affected proteins in mouse models of disease and other live animal test settings.
Keywords: Alzheimer's disease; SILAM; aging; neurodegeneration; protein turnover; proteomics.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare no competing interests.
Figures
References
-
- Dwyer B.E., Fando J.L., Wasterlain C.G. Rat brain protein synthesis declines during postdevelopmental aging. J. Neurochem. 1980;35:746–749. - PubMed
-
- Fornasiero E.F., Mandad S., Wildhagen H., Alevra M., Rammner B., Keihani S., Opazo F., Urban I., Ischebeck T., Sakib M.S., Fard M.K., Kirli K., Centeno T.P., Vidal R.O., Rahman R.-U., et al. Precisely measured protein lifetimes in the mouse brain reveal differences across tissues and subcellular fractions. Nat. Commun. 2018;9:4230. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
