

Isoform-dependent lysosomal degradation and internalization of apolipoprotein E requires autophagy proteins
- PMID: 34982109
- PMCID: PMC8917355
- DOI: 10.1242/jcs.258687
Isoform-dependent lysosomal degradation and internalization of apolipoprotein E requires autophagy proteins
Abstract
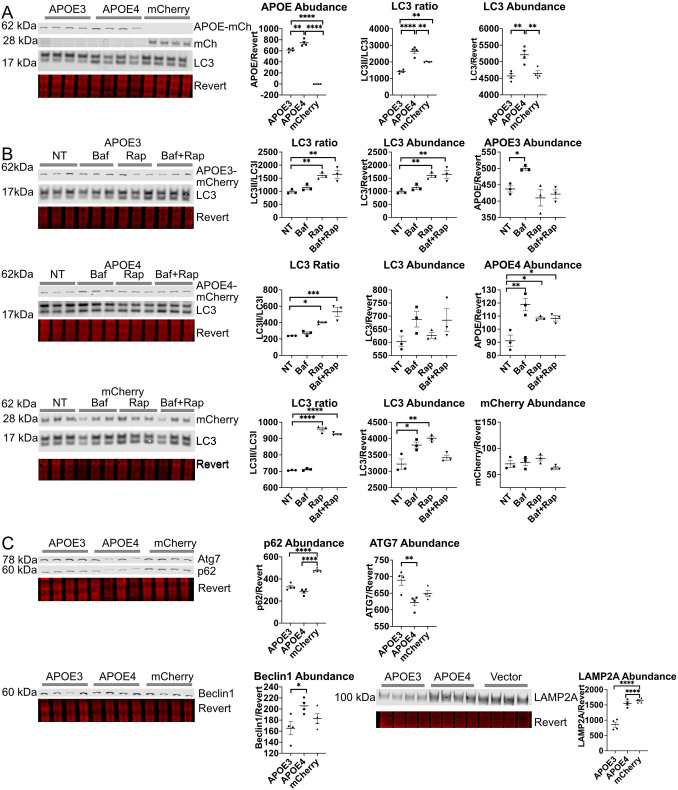
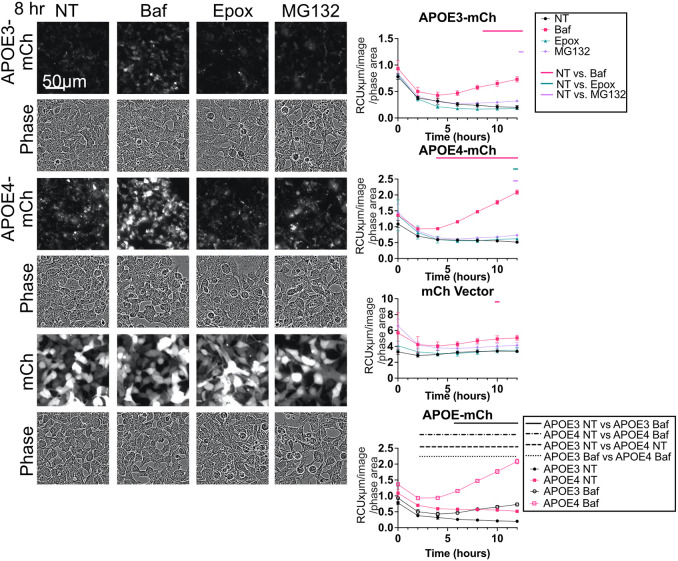
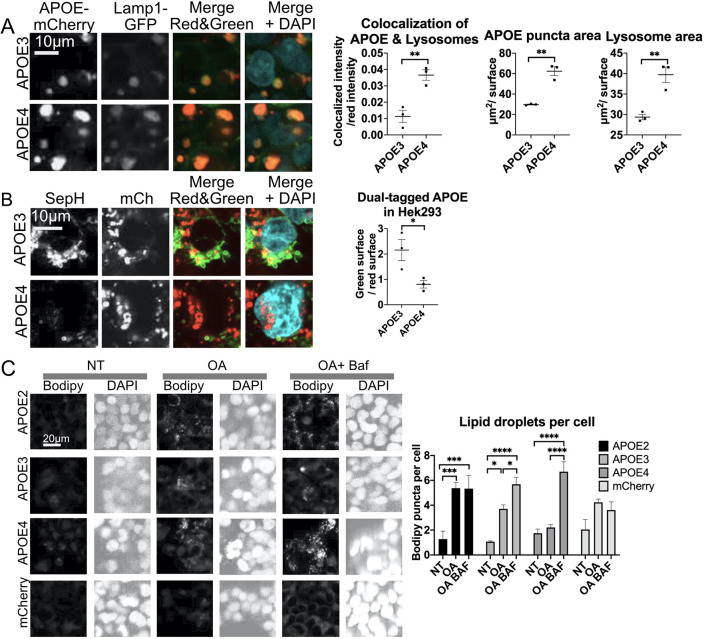
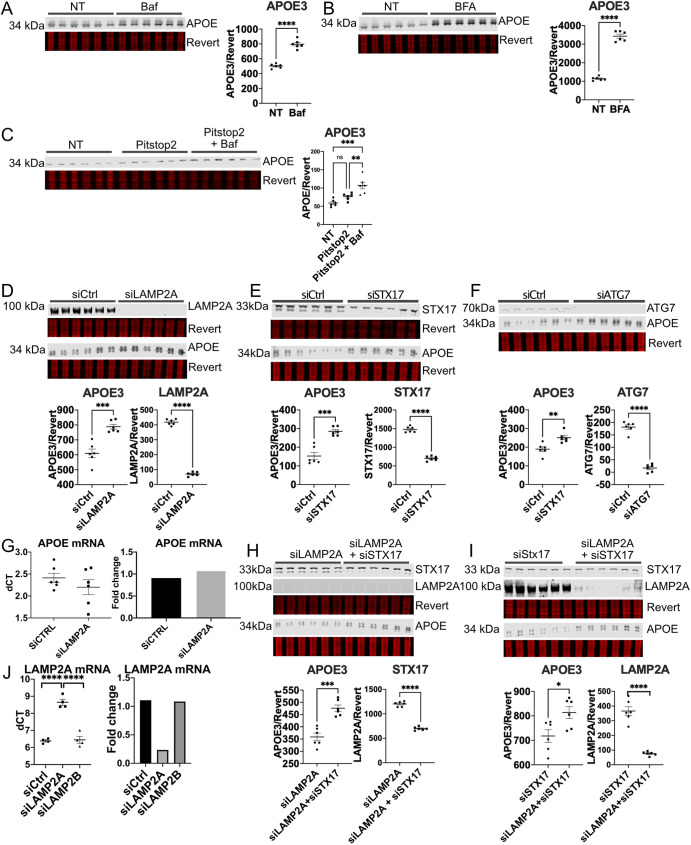
The human apolipoprotein E4 isoform (APOE4) is the strongest genetic risk factor for late-onset Alzheimer's disease (AD), and lysosomal dysfunction has been implicated in AD pathogenesis. We found, by examining cells stably expressing each APOE isoform, that APOE4 increases lysosomal trafficking, accumulates in enlarged lysosomes and late endosomes, alters autophagic flux and the abundance of autophagy proteins and lipid droplets, and alters the proteomic contents of lysosomes following internalization. We investigated APOE-related lysosomal trafficking further in cell culture, and found that APOE from the post-Golgi compartment is degraded through autophagy. We found that this autophagic process requires the lysosomal membrane protein LAMP2 in immortalized neuron-like and hepatic cells, and in mouse brain tissue. Several macroautophagy-associated proteins were also required for autophagic degradation and internalization of APOE in hepatic cells. The dysregulated autophagic flux and lysosomal trafficking of APOE4 that we observed suggest a possible novel mechanism that might contribute to AD pathogenesis. This article has an associated First Person interview with the first author of the paper.
Keywords: APOE; APOE4; Alzheimer's disease; Chaperone-mediated autophagy; LC3-associated endocytosis.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
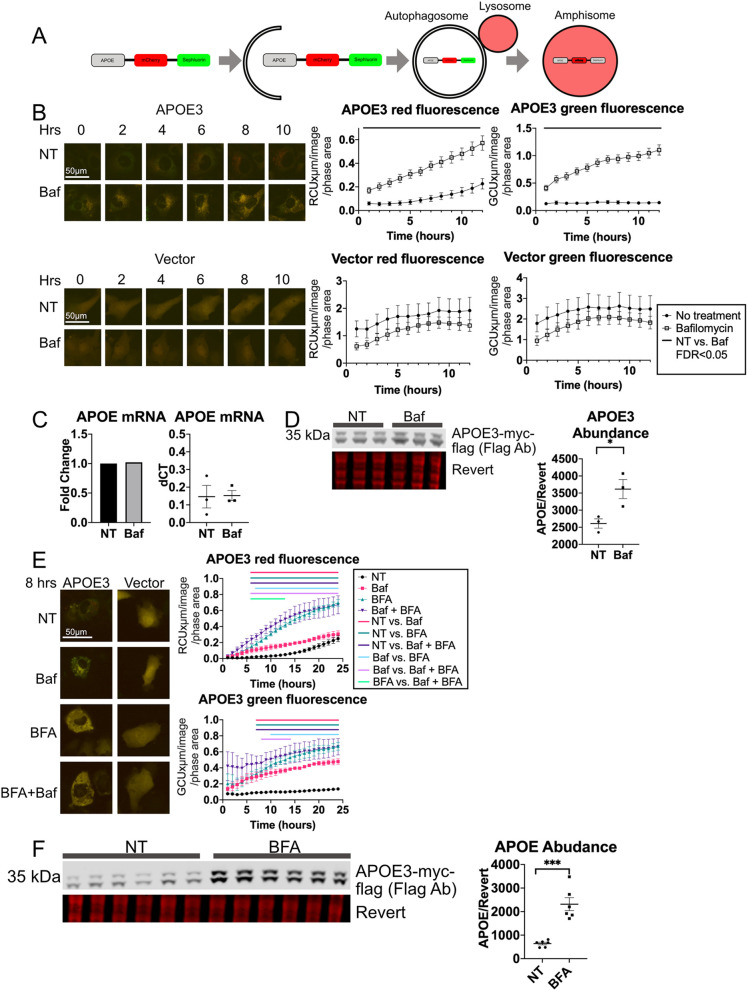
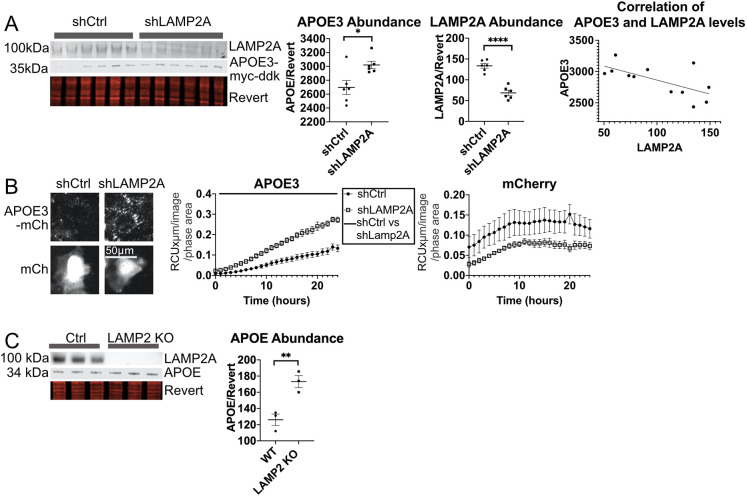
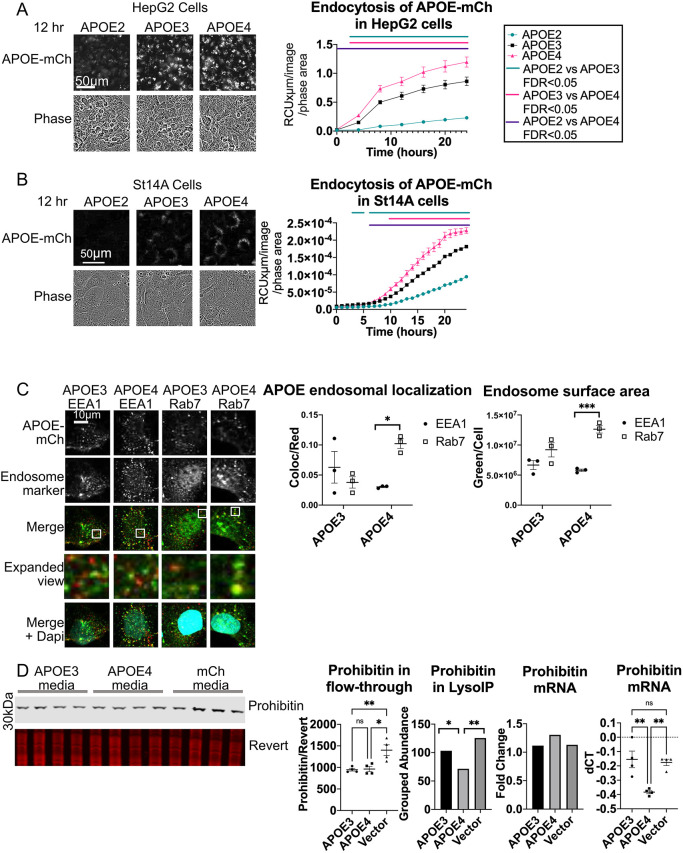
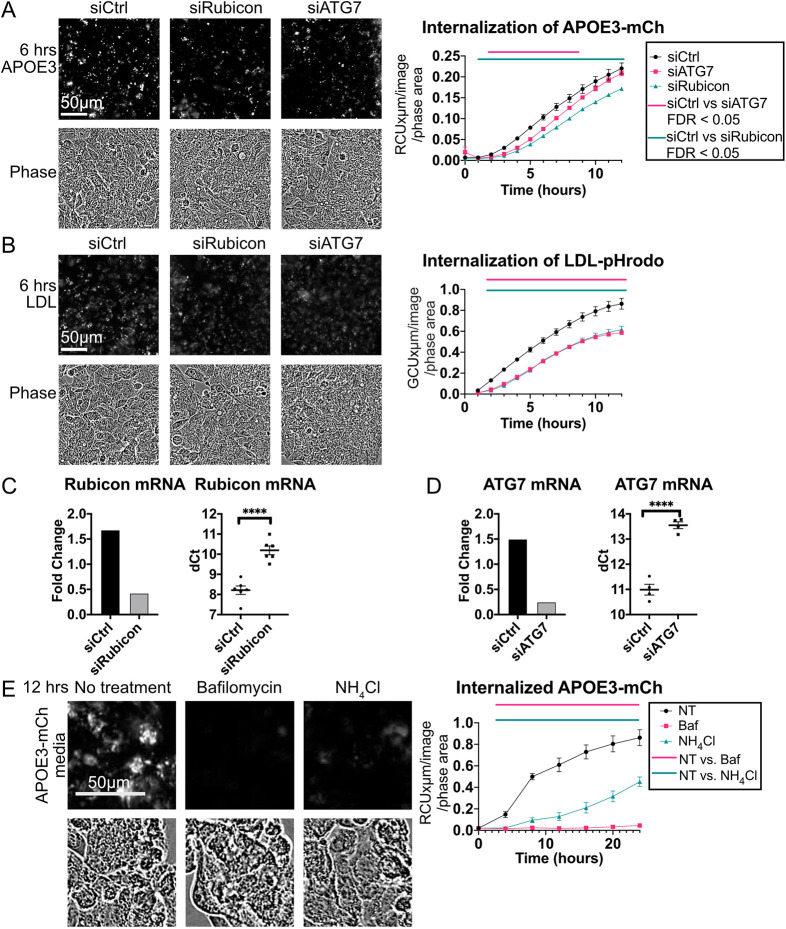
References
-
- Abu-Remaileh, M., Wyant, G. A., Kim, C., Laqtom, N. N., Abbasi, M., Chan, S. H., Freinkman, E. and Sabatini, D. M. (2017). Lysosomal metabolomics reveals V-ATPase- and mTOR-dependent regulation of amino acid efflux from lysosomes. Science (80-.) 358, 807-813. 10.1126/science.aan6298 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 1F30AG060704-01A1/NH/NIH HHS/United States
- T32 GM008620/GM/NIGMS NIH HHS/United States
- NS116872/NH/NIH HHS/United States
- R35 NS116872/NS/NINDS NIH HHS/United States
- P50 AG016573/AG/NIA NIH HHS/United States
- R56 NS072453/NS/NINDS NIH HHS/United States
- T32 AG000096/AG/NIA NIH HHS/United States
- F30 AG060704/AG/NIA NIH HHS/United States
- S10 OD010669/OD/NIH HHS/United States
- 1F30AG060704/NH/NIH HHS/United States
- R01 NS090390/NS/NINDS NIH HHS/United States
- R56 NS090390/NS/NINDS NIH HHS/United States
- R01 NS072453/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
