

Lipid-Protein Interactions in Plasma Membrane Organization and Function
- PMID: 34982570
- PMCID: PMC12101515
- DOI: 10.1146/annurev-biophys-090721-072718
Lipid-Protein Interactions in Plasma Membrane Organization and Function
Abstract
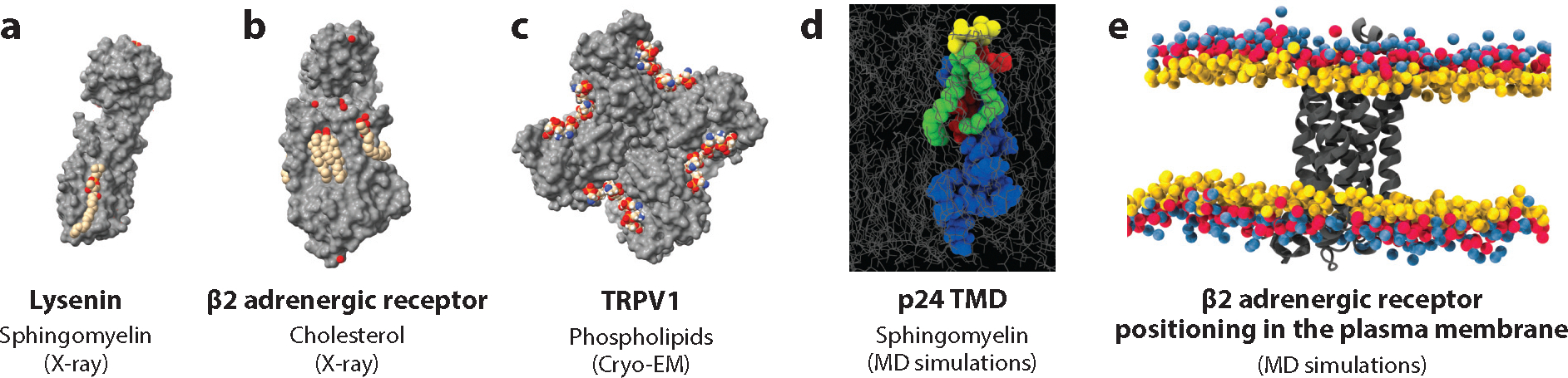
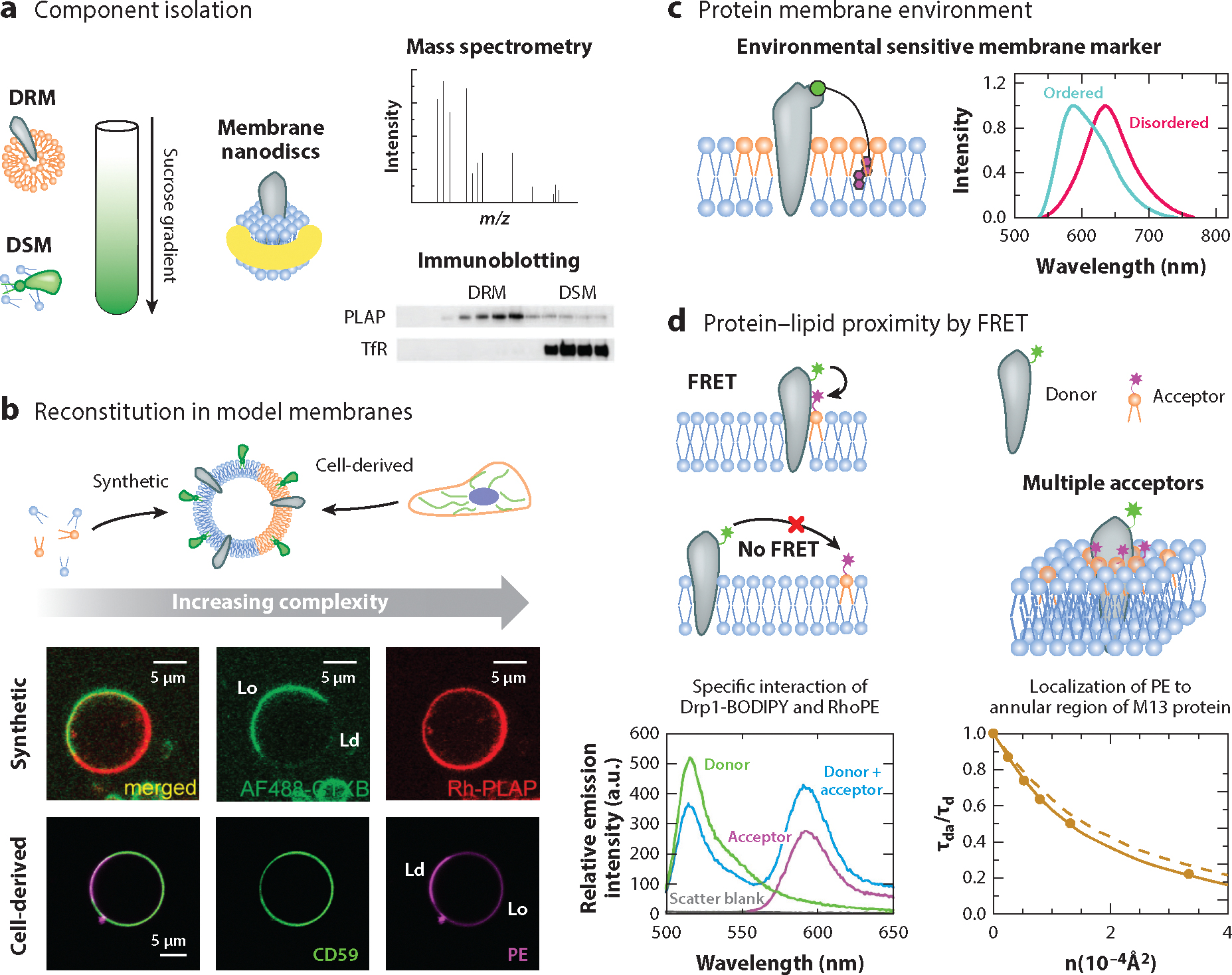
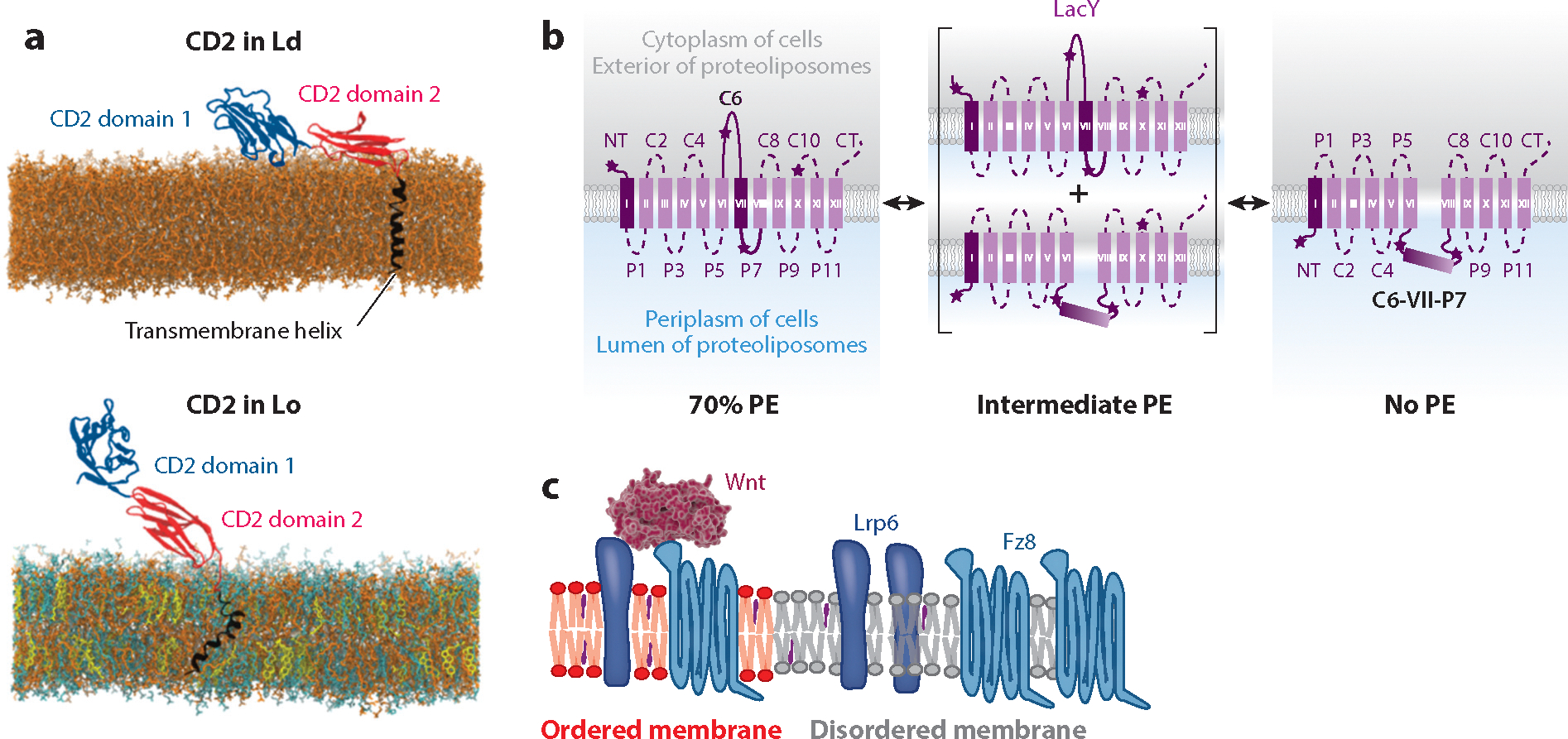
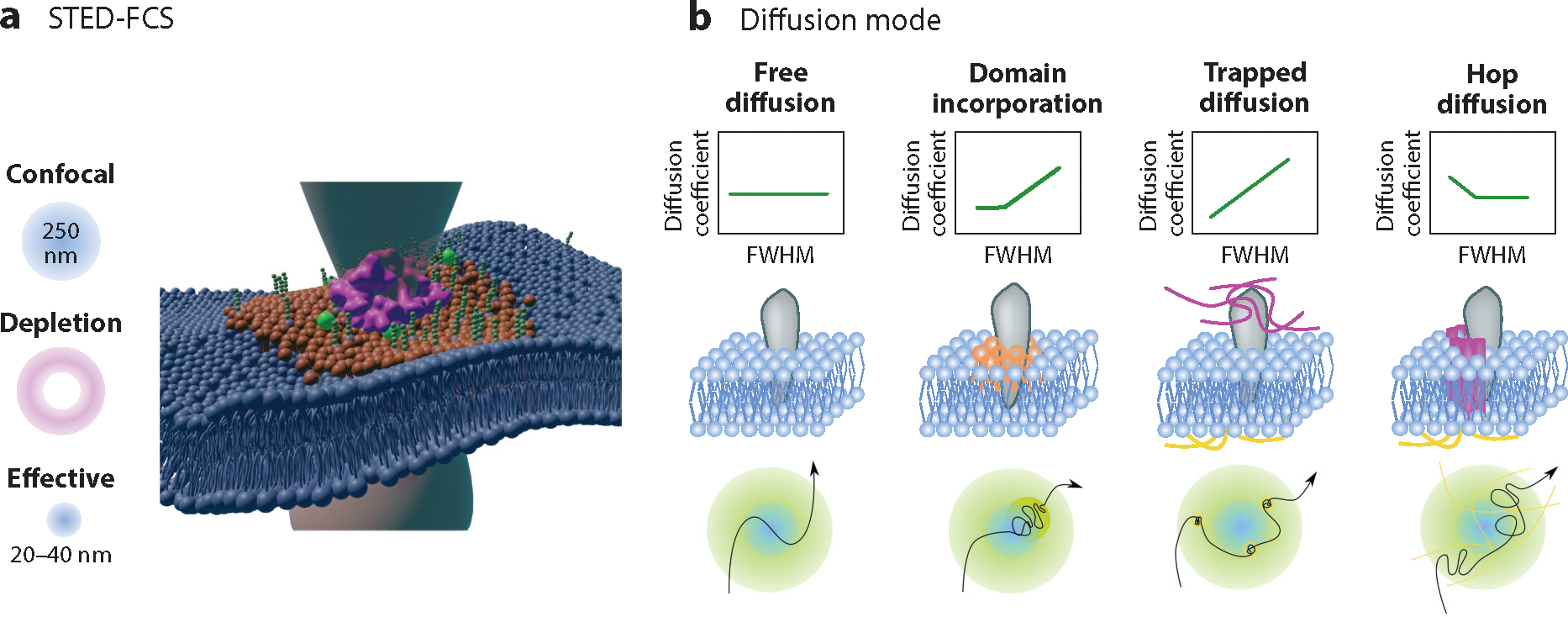
Lipid-protein interactions in cells are involved in various biological processes, including metabolism, trafficking, signaling, host-pathogen interactions, and transmembrane transport. At the plasma membrane, lipid-protein interactions play major roles in membrane organization and function. Several membrane proteins have motifs for specific lipid binding, which modulate protein conformation and consequent function. In addition to such specific lipid-protein interactions, protein function can be regulated by the dynamic, collective behavior of lipids in membranes. Emerging analytical, biochemical, and computational technologies allow us to study the influence of specific lipid-protein interactions, as well as the collective behavior of membranes on protein function. In this article, we review the recent literature on lipid-protein interactions with a specific focus on the current state-of-the-art technologies that enable novel insights into these interactions.
Keywords: collective membrane properties; cryo-EM; lipid-binding motifs; membrane biophysics; molecular dynamics; super-resolution imaging.
Figures
Similar articles
-
The interactions of peripheral membrane proteins with biological membranes.Chem Phys Lipids. 2015 Nov;192:51-59. doi: 10.1016/j.chemphyslip.2015.07.015. Epub 2015 Jul 29. Chem Phys Lipids. 2015. PMID: 26232665 Review.
-
Membrane-protein interactions in cell signaling and membrane trafficking.Annu Rev Biophys Biomol Struct. 2005;34:119-51. doi: 10.1146/annurev.biophys.33.110502.133337. Annu Rev Biophys Biomol Struct. 2005. PMID: 15869386 Review.
-
Emerging Diversity in Lipid-Protein Interactions.Chem Rev. 2019 May 8;119(9):5775-5848. doi: 10.1021/acs.chemrev.8b00451. Epub 2019 Feb 13. Chem Rev. 2019. PMID: 30758191 Free PMC article. Review.
-
A simple guide to biochemical approaches for analyzing protein-lipid interactions.Mol Biol Cell. 2012 Aug;23(15):2823-30. doi: 10.1091/mbc.E11-07-0645. Mol Biol Cell. 2012. PMID: 22848065 Free PMC article. Review.
-
The importance of membrane defects-lessons from simulations.Acc Chem Res. 2014 Aug 19;47(8):2244-51. doi: 10.1021/ar4002729. Epub 2014 Jun 3. Acc Chem Res. 2014. PMID: 24892900
Cited by
-
Deciphering Lipid Arrangement in Phosphatidylserine/Phosphatidylcholine Mixed Membranes: Simulations and Experiments.Langmuir. 2023 Dec 26;39(51):18995-19007. doi: 10.1021/acs.langmuir.3c03061. Epub 2023 Dec 14. Langmuir. 2023. PMID: 38096496 Free PMC article.
-
The Myth of The Annular Lipids.Biomedicines. 2022 Oct 22;10(11):2672. doi: 10.3390/biomedicines10112672. Biomedicines. 2022. PMID: 36359192 Free PMC article. Review.
-
Identification of Protein Content of Immature Canine Oocytes.Vet Med Sci. 2025 Mar;11(2):e70247. doi: 10.1002/vms3.70247. Vet Med Sci. 2025. PMID: 39992178 Free PMC article.
-
Regulation of membrane protein structure and function by their lipid nano-environment.Nat Rev Mol Cell Biol. 2023 Feb;24(2):107-122. doi: 10.1038/s41580-022-00524-4. Epub 2022 Sep 2. Nat Rev Mol Cell Biol. 2023. PMID: 36056103 Free PMC article. Review.
-
GLUT4 dispersal at the plasma membrane of adipocytes: a super-resolved journey.Biosci Rep. 2023 Oct 31;43(10):BSR20230946. doi: 10.1042/BSR20230946. Biosci Rep. 2023. PMID: 37791639 Free PMC article. Review.
References
-
- Bacia K, Schuette CG, Kahya N, Jahn R, Schwille P. 2004. SNAREs prefer liquid-disordered over “raft” (liquid-ordered) domains when reconstituted into giant unilamellar vesicles. J. Biol. Chem. 279(36):37951–55 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
