

Inactivating Celsr2 promotes motor axon fasciculation and regeneration in mouse and human
- PMID: 34983065
- PMCID: PMC9014747
- DOI: 10.1093/brain/awab317
Inactivating Celsr2 promotes motor axon fasciculation and regeneration in mouse and human
Abstract
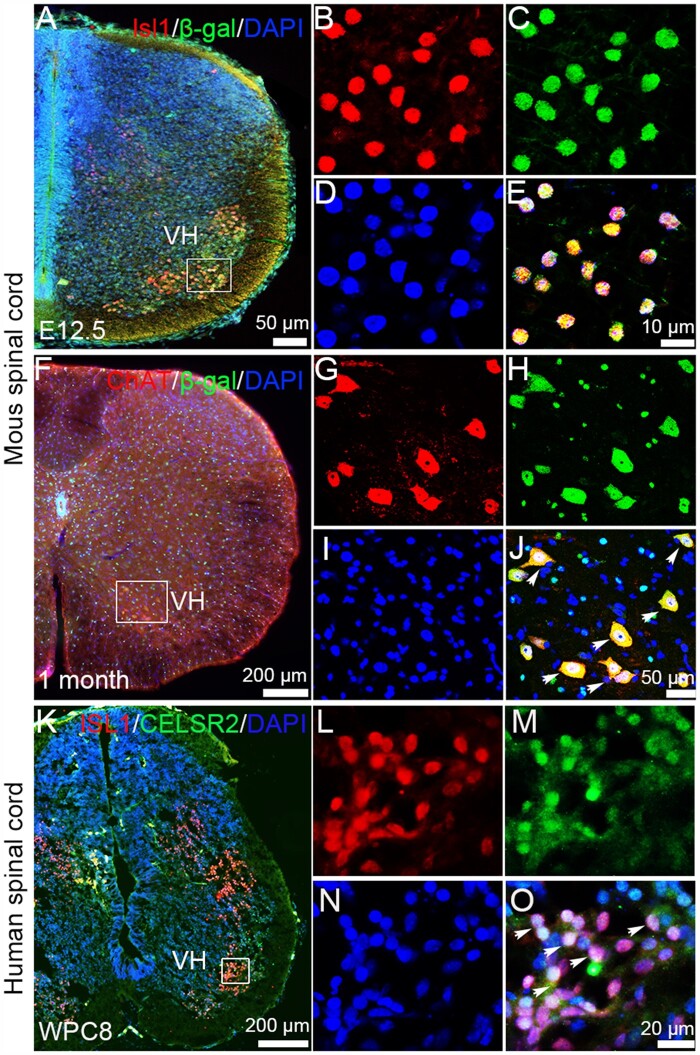
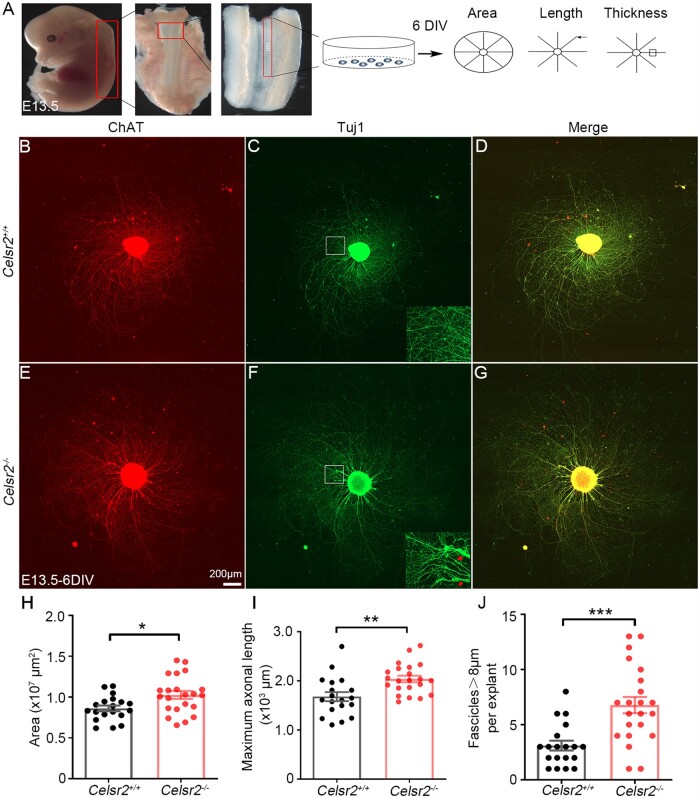
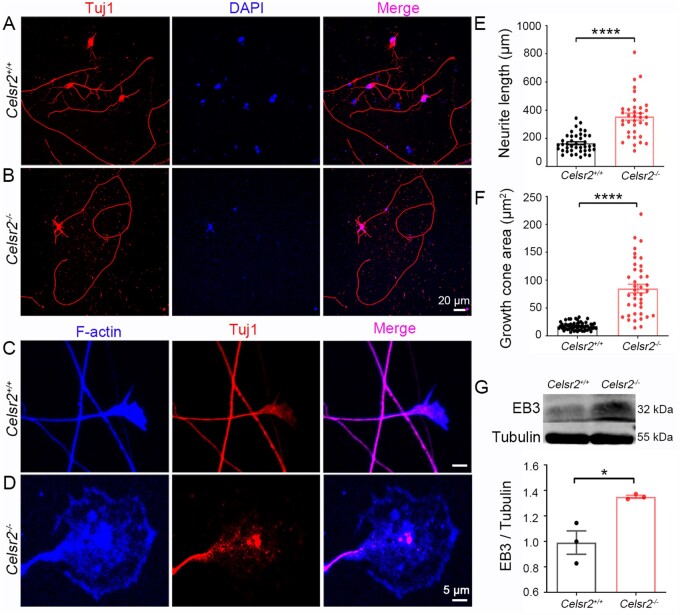
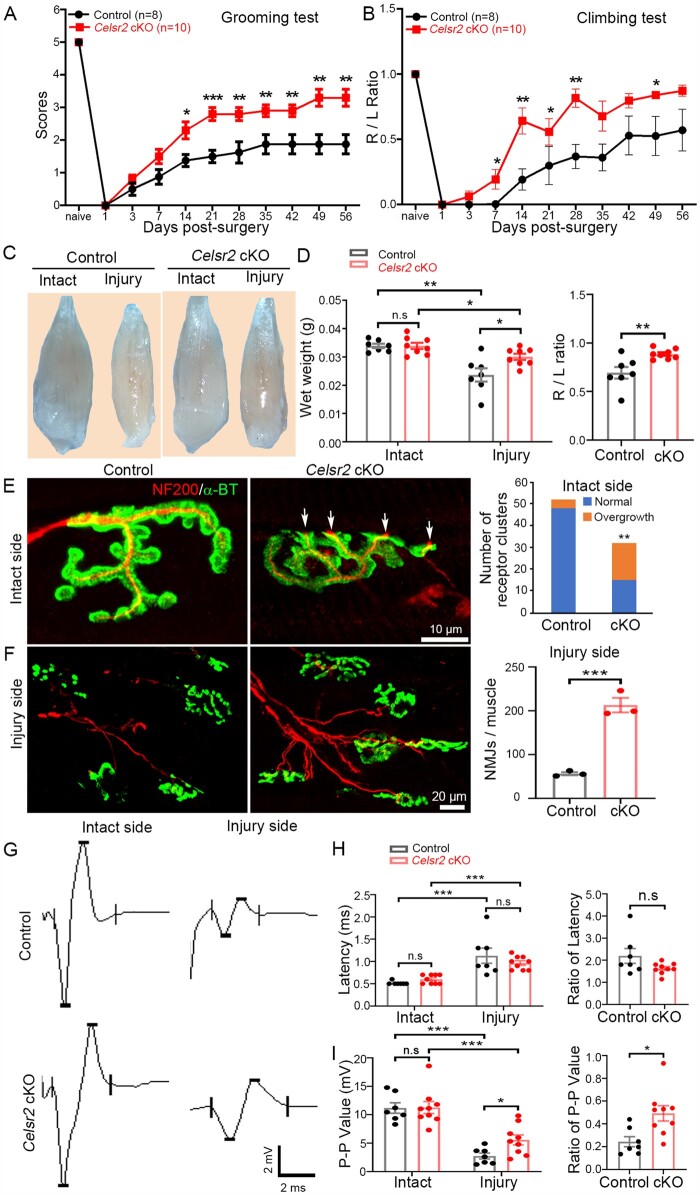
Understanding new modulators of axon regeneration is central to neural repair. Our previous work demonstrated critical roles of atypical cadherin Celsr2 during neural development, including cilia organization, neuron migration and axon navigation. Here, we address its role in axon regeneration. We show that Celsr2 is highly expressed in both mouse and human spinal motor neurons. Celsr2 knockout promotes axon regeneration and fasciculation in mouse cultured spinal explants. Similarly, cultured Celsr2 mutant motor neurons extend longer neurites and larger growth cones, with increased expression of end-binding protein 3 and higher potassium-induced calcium influx. Mice with Celsr2 conditional knockout in spinal motor neurons do not exhibit any behavioural deficits; however, after branchial plexus injury, axon regeneration and functional forelimb locomotor recovery are significantly improved. Similarly, knockdown of CELSR2 using shRNA interference in cultured human spinal motor explants and motor neurons increases axonal fasciculation and growth. In mouse adult spinal cord after root avulsion, in mouse embryonic spinal cords, and in cultured human motor neurons, Celsr2 downregulation is accompanied by increased levels of GTP-bound Rac1 and Cdc42, and of JNK and c-Jun. In conclusion, Celsr2 negatively regulates motor axon regeneration and is a potential target to improve neural repair.
Keywords: axon regeneration; brachial plexus injury; human embryos; root avulsion; spinal motor neurons.
© The Author(s) (2022). Published by Oxford University Press on behalf of the Guarantors of Brain.
Figures
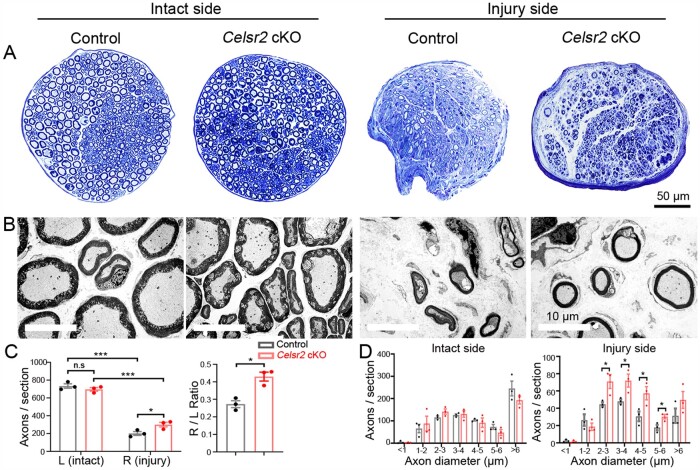
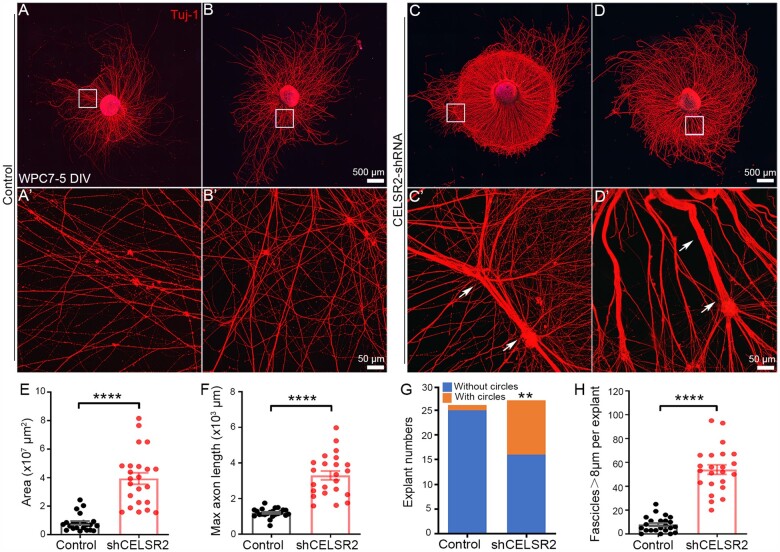
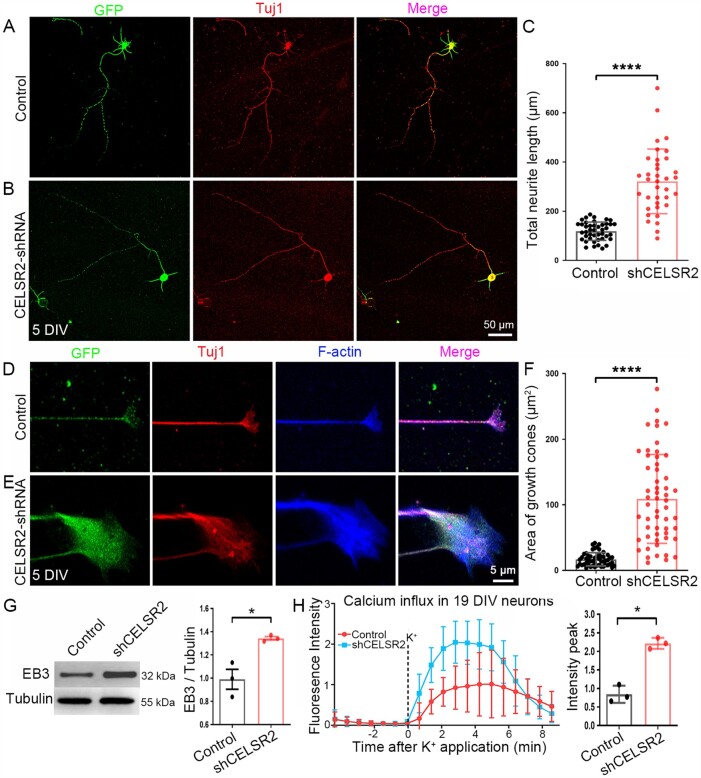
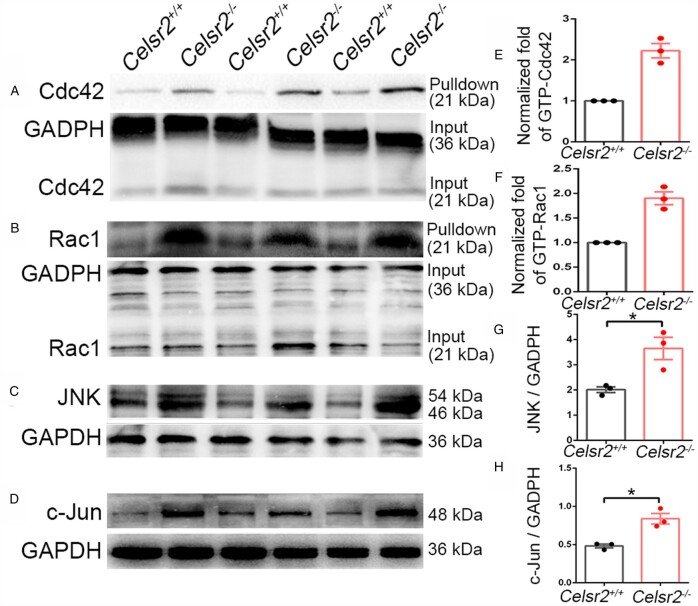
Comment in
-
CELSR2, a new player in motor neuron axon growth and regeneration.Brain. 2022 Apr 18;145(2):420-422. doi: 10.1093/brain/awac021. Brain. 2022. PMID: 35259228 No abstract available.
References
-
- Liu K, Tedeschi A, Park KK, He Z.. Neuronal intrinsic mechanisms of axon regeneration. Annu Rev Neurosci. 2011;34:131–152. - PubMed
-
- Cullheim S, Carlstedt T, Linda H, Risling M, Ulfhake B.. Motoneurons reinnervate skeletal muscle after ventral root implantation into the spinal cord of the cat. Neuroscience. 1989;29(3):725–733. - PubMed
-
- Carlstedt T, Grane P, Hallin RG, Noren G.. Return of function after spinal cord implantation of avulsed spinal nerve roots. Lancet. 1995;346(8986):1323–1325. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
