

PPARα and PPARγ activation is associated with pleural mesothelioma invasion but therapeutic inhibition is ineffective
- PMID: 34984327
- PMCID: PMC8692993
- DOI: 10.1016/j.isci.2021.103571
PPARα and PPARγ activation is associated with pleural mesothelioma invasion but therapeutic inhibition is ineffective
Abstract
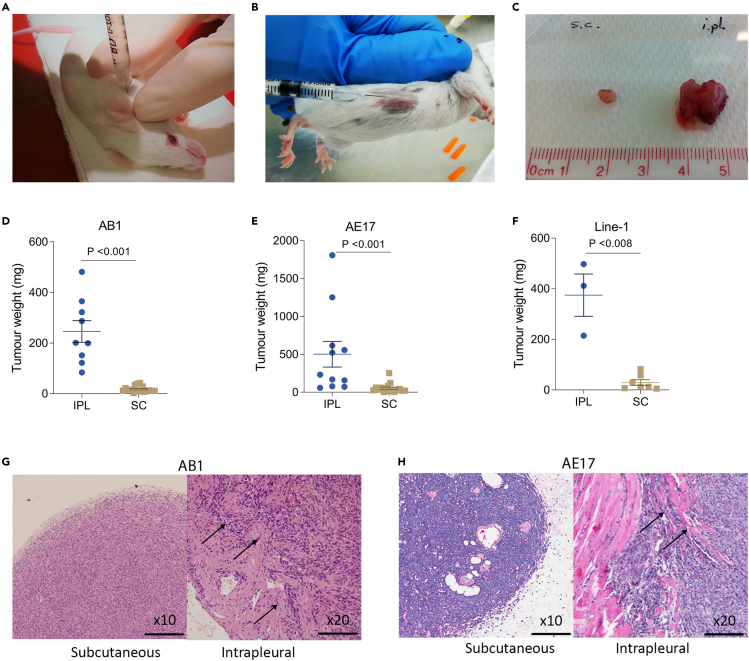
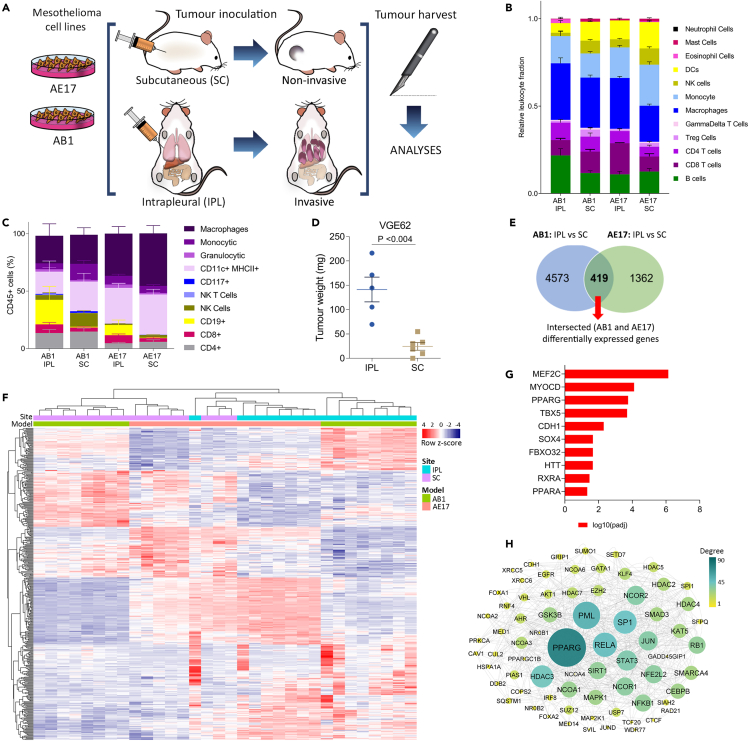
Mesothelioma is a cancer that typically originates in the pleura of the lungs. It rapidly invades the surrounding tissues, causing pain and shortness of breath. We compared cell lines injected either subcutaneously or intrapleurally and found that only the latter resulted in invasive and rapid growth. Pleural tumors displayed a transcriptional signature consistent with increased activity of nuclear receptors PPARα and PPARγ and with an increased abundance of endogenous PPAR-activating ligands. We found that chemical probe GW6471 is a potent, dual PPARα/γ antagonist with anti-invasive and anti-proliferative activity in vitro. However, administration of GW6471 at doses that provided sustained plasma exposure levels sufficient for inhibition of PPARα/γ transcriptional activity did not result in significant anti-mesothelioma activity in mice. Lastly, we demonstrate that the in vitro anti-tumor effect of GW6471 is off-target. We conclude that dual PPARα/γ antagonism alone is not a viable treatment modality for mesothelioma.
Keywords: Biological sciences; Metabolomics; Transcriptomics.
© 2021 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Andrews, S., 2010. FASTQC. A quality control tool for high throughput sequence data.
-
- Aota S., Nagai T., Olden K., Akiyama S.K., Yamada K.M. Fibronectin and integrins in cell adhesion, signaling, and morphogenesis. Biochem. Soc. Trans. 1991;19:830–835. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
