

Hippocampal acetylcholine modulates stress-related behaviors independent of specific cholinergic inputs
- PMID: 34997190
- PMCID: PMC9106825
- DOI: 10.1038/s41380-021-01404-7
Hippocampal acetylcholine modulates stress-related behaviors independent of specific cholinergic inputs
Abstract
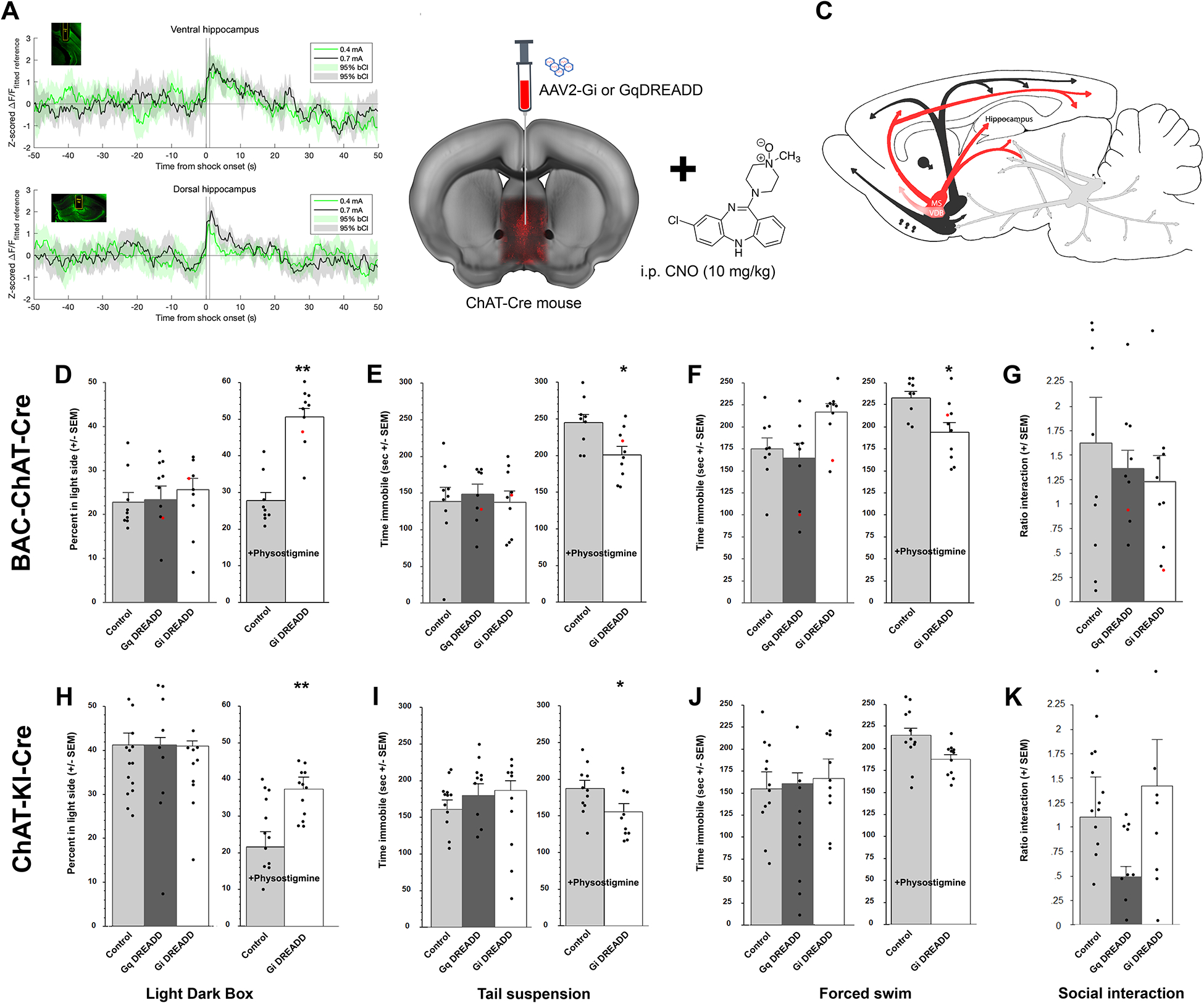
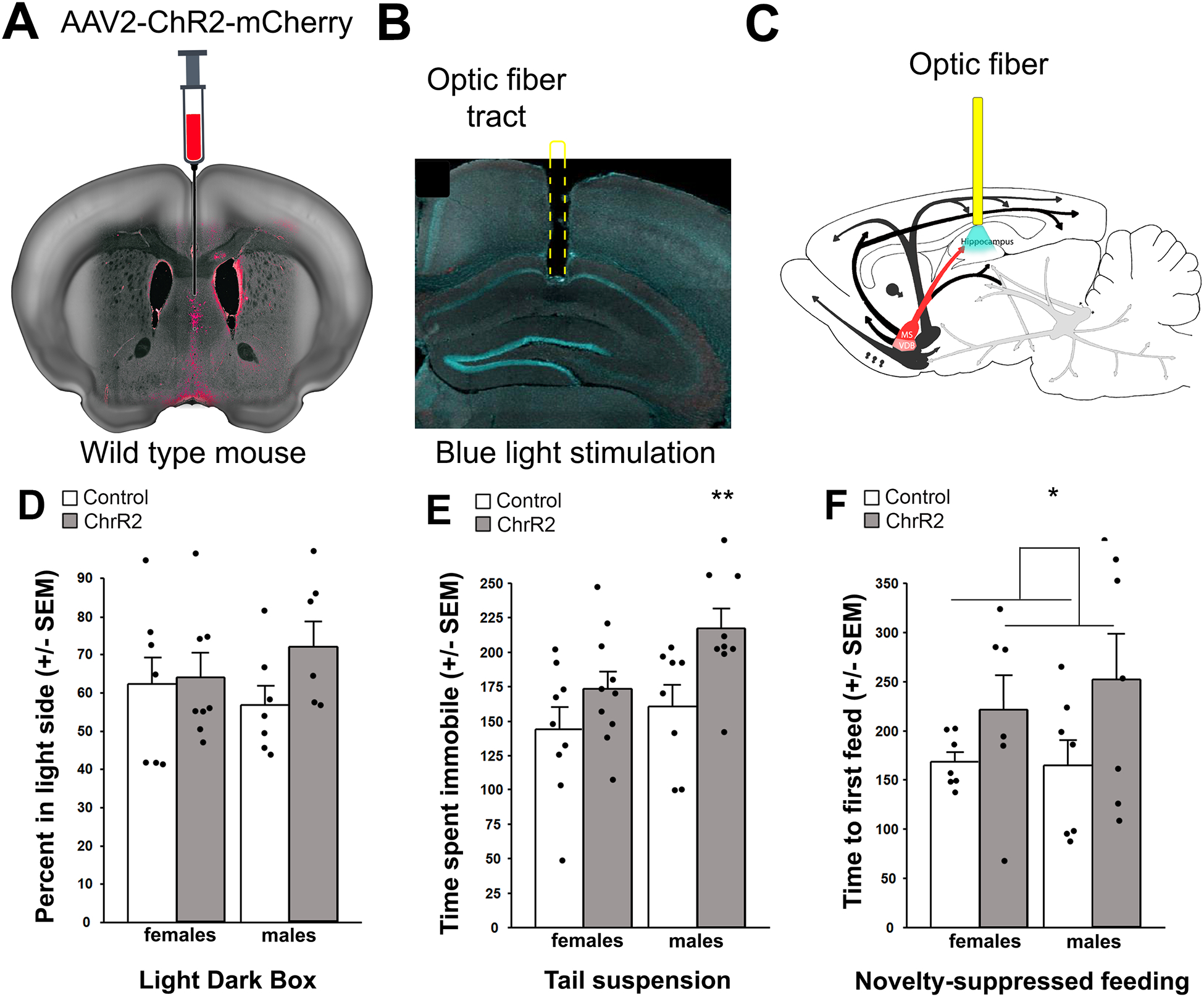
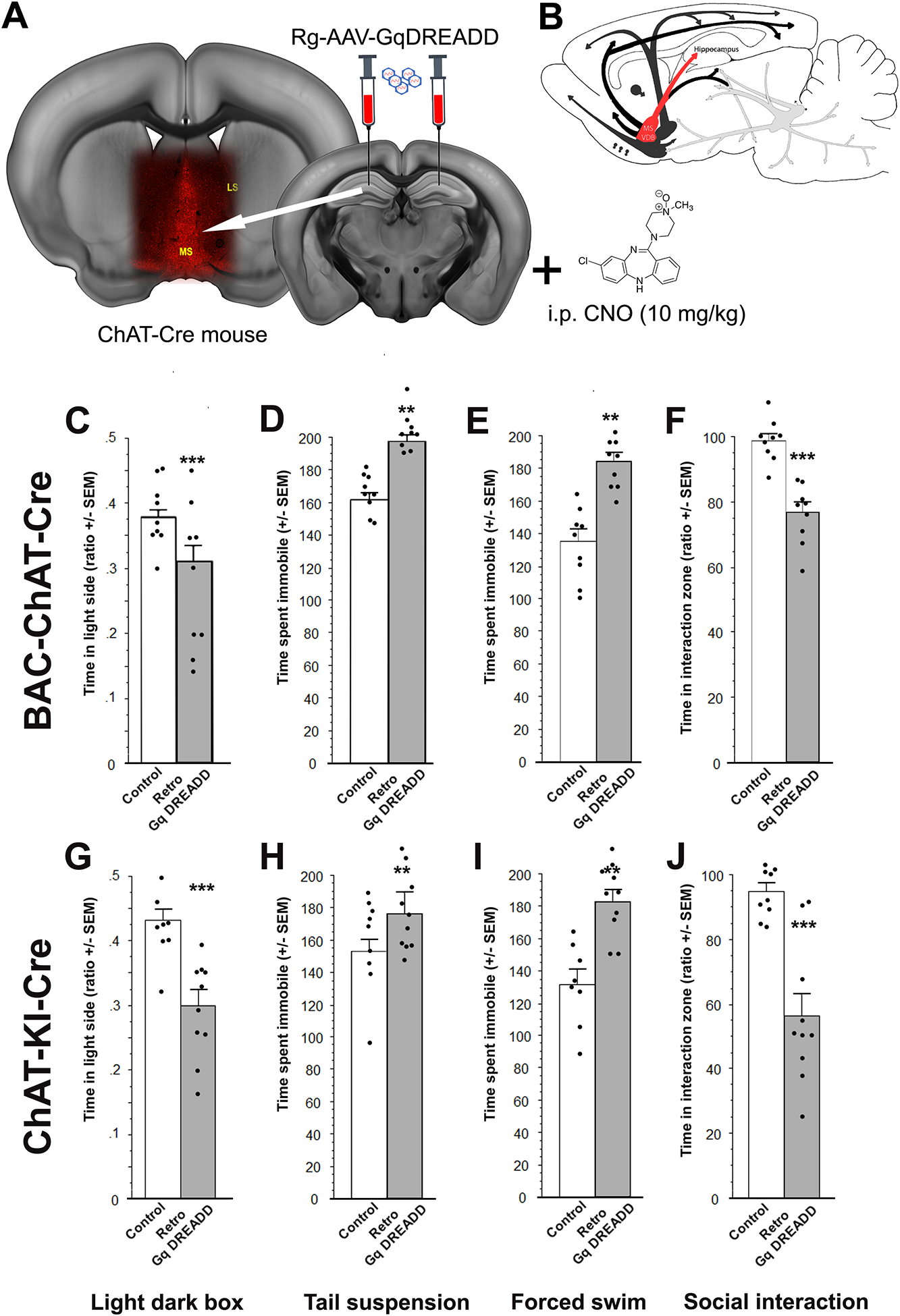
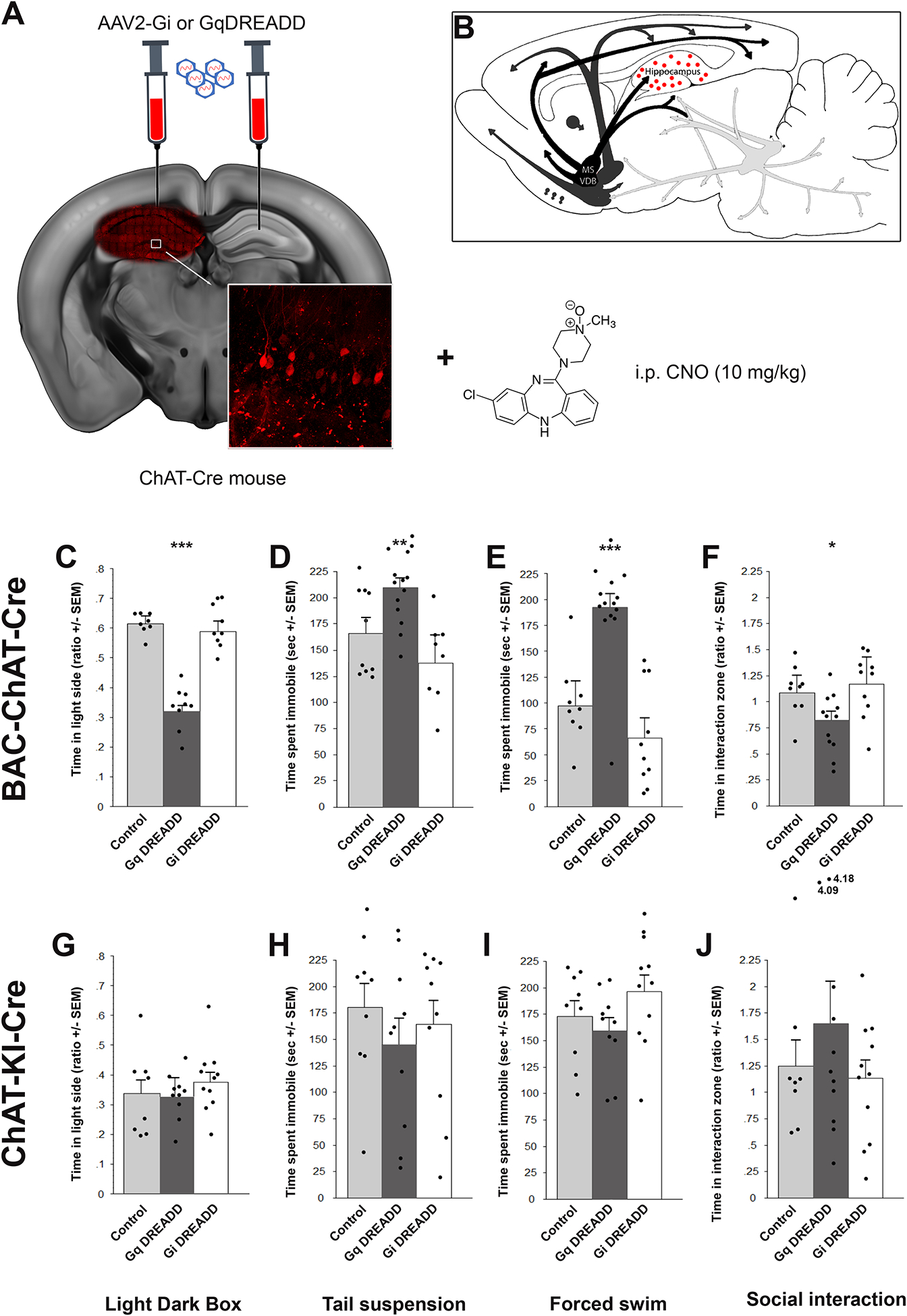
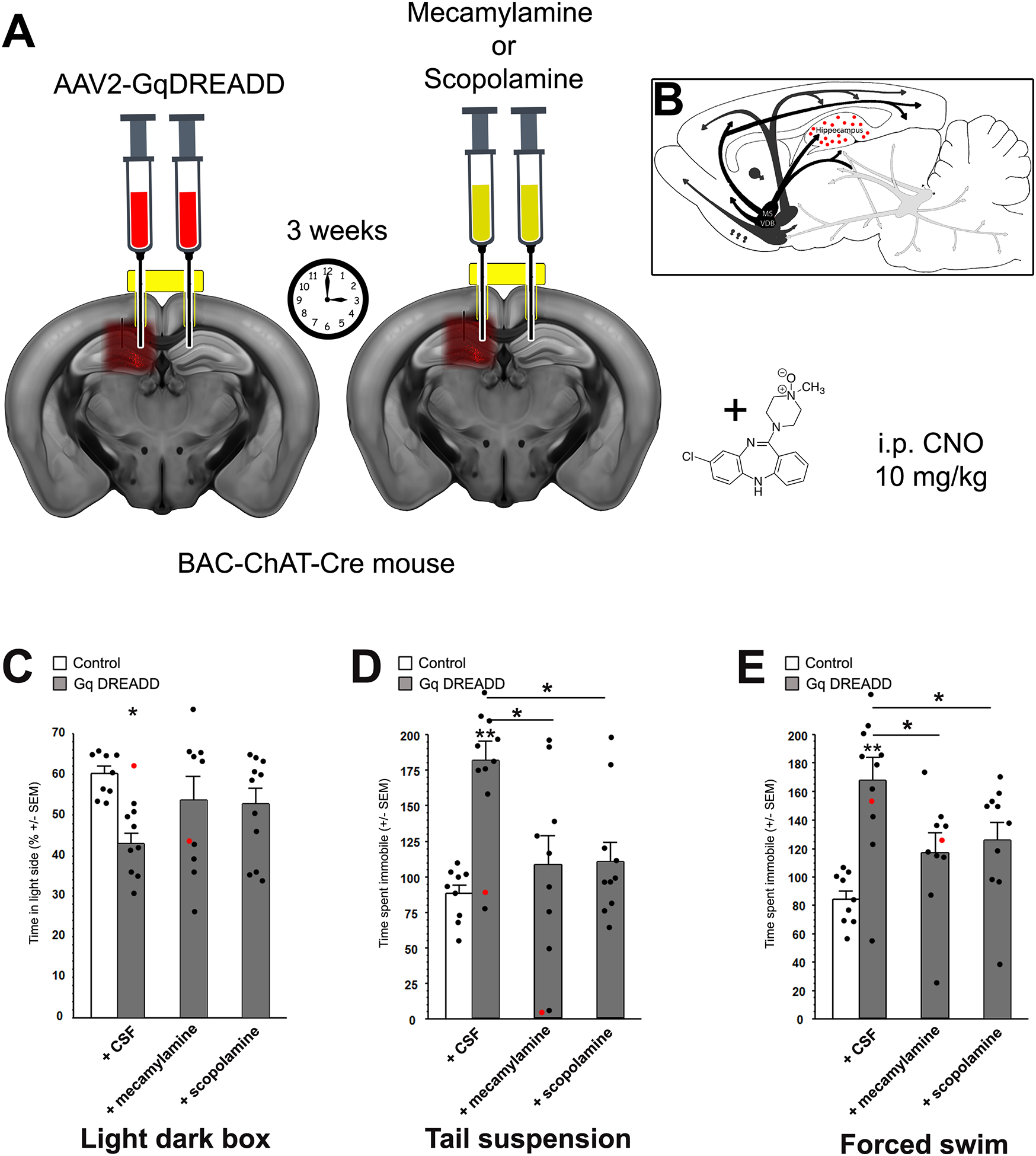
Acetylcholine (ACh) levels are elevated in actively depressed subjects. Conversely, antagonism of either nicotinic or muscarinic ACh receptors can have antidepressant effects in humans and decrease stress-relevant behaviors in rodents. Consistent with a role for ACh in mediating maladaptive responses to stress, brain ACh levels increase in response to stressful challenges, whereas systemically blocking acetylcholinesterase (AChE, the primary ACh degradative enzyme) elicits depression-like symptoms in human subjects, and selectively blocking AChE in the hippocampus increases relevant behaviors in rodents. We used an ACh sensor to characterize stress-evoked ACh release, then used chemogenetic, optogenetic and pharmacological approaches to determine whether cholinergic inputs from the medial septum/diagonal bands of Broca (MSDBB) or ChAT-positive neurons intrinsic to the hippocampus mediate stress-relevant behaviors in mice. Chemogenetic inhibition or activation of MSDBB cholinergic neurons did not result in significant behavioral effects, while inhibition attenuated the behavioral effects of physostigmine. In contrast, optogenetic stimulation of septohippocampal terminals or selective chemogenetic activation of ChAT-positive inputs to hippocampus increased stress-related behaviors. Finally, stimulation of sparse ChAT-positive hippocampal neurons increased stress-related behaviors in one ChAT-Cre line, which were attenuated by local infusion of cholinergic antagonists. These studies suggest that ACh signaling results in maladaptive behavioral responses to stress if the balance of signaling is shifted toward increased hippocampal engagement.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
CONFLICT OF INTEREST
None
Figures
References
-
- Janowsky DS, el-Yousef MK, Davis JM, Sekerke HJ. A cholinergic-adrenergic hypothesis of mania and depression. Lancet 1972; 2(7778): 632–635. - PubMed
-
- Risch SC, Cohen RM, Janowsky DS, Kalin NH, Sitaram N, Gillin JC et al. Physostigmine induction of depressive symptomatology in normal human subjects. Psychiatry research 1981; 4(1): 89–94. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
