

Chloroplast Ribosomes Interact With the Insertase Alb3 in the Thylakoid Membrane
- PMID: 35003166
- PMCID: PMC8733628
- DOI: 10.3389/fpls.2021.781857
Chloroplast Ribosomes Interact With the Insertase Alb3 in the Thylakoid Membrane
Abstract
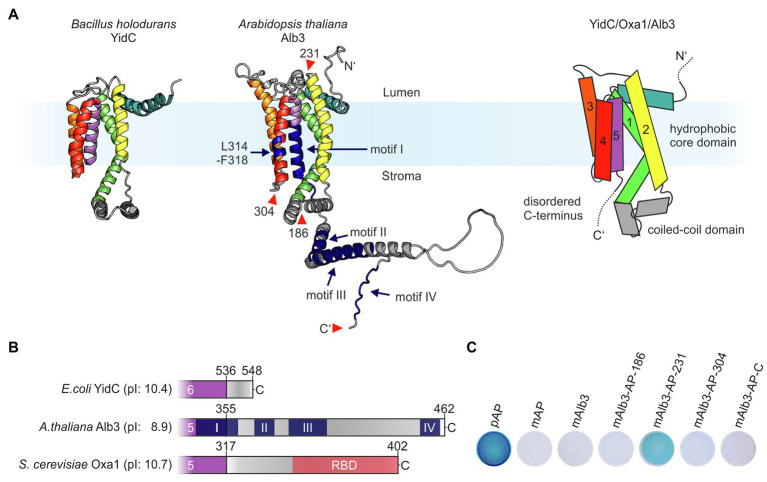
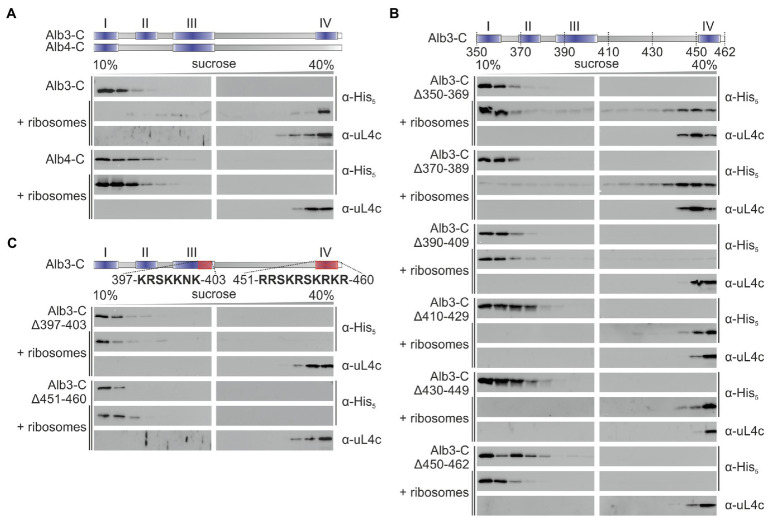
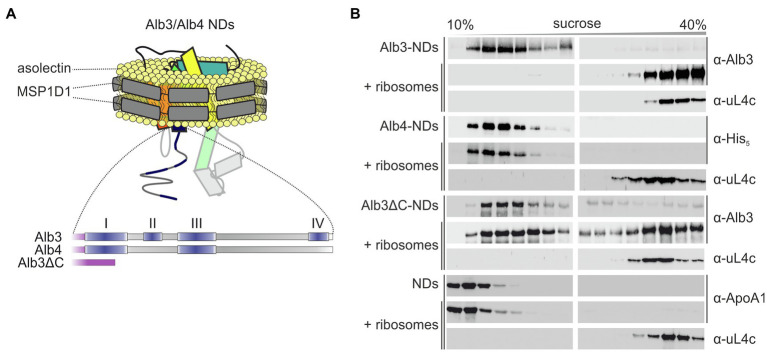
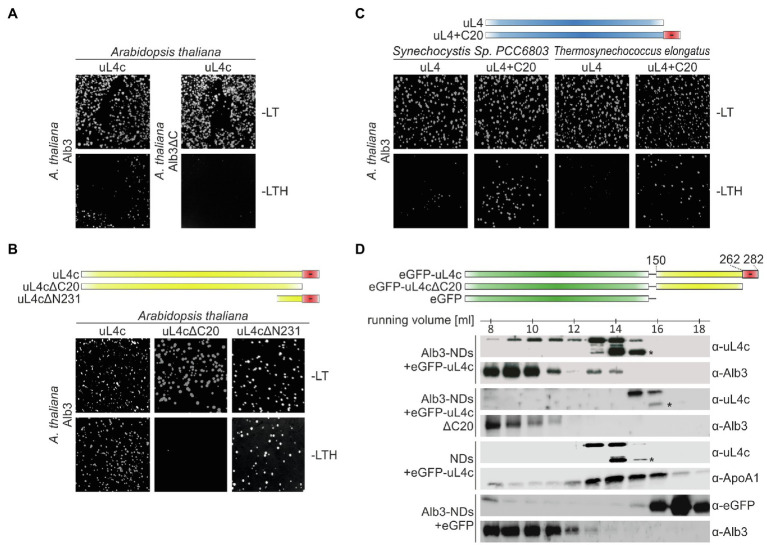
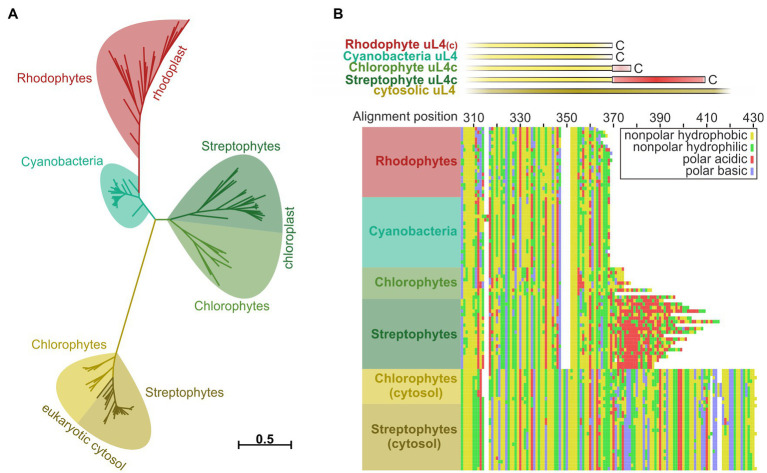
Members of the Oxa1/YidC/Alb3 protein family are involved in the insertion, folding, and assembly of membrane proteins in mitochondria, bacteria, and chloroplasts. The thylakoid membrane protein Alb3 mediates the chloroplast signal recognition particle (cpSRP)-dependent posttranslational insertion of nuclear-encoded light harvesting chlorophyll a/b-binding proteins and participates in the biogenesis of plastid-encoded subunits of the photosynthetic complexes. These subunits are cotranslationally inserted into the thylakoid membrane, yet very little is known about the molecular mechanisms underlying docking of the ribosome-nascent chain complexes to the chloroplast SecY/Alb3 insertion machinery. Here, we show that nanodisc-embedded Alb3 interacts with ribosomes, while the homolog Alb4, also located in the thylakoid membrane, shows no ribosome binding. Alb3 contacts the ribosome with its C-terminal region and at least one additional binding site within its hydrophobic core region. Within the C-terminal region, two conserved motifs (motifs III and IV) are cooperatively required to enable the ribosome contact. Furthermore, our data suggest that the negatively charged C-terminus of the ribosomal subunit uL4c is involved in Alb3 binding. Phylogenetic analyses of uL4 demonstrate that this region newly evolved in the green lineage during the transition from aquatic to terrestrial life.
Keywords: Oxa1/YidC/Alb3 protein family; cotranslational protein transport; nanodiscs; ribosomes; thylakoid membrane biogenesis.
Copyright © 2021 Ackermann, Dünschede, Pietzenuk, Justesen, Krämer, Hofmann, Günther Pomorski and Schünemann.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources