

Colonies of the marine cyanobacterium Trichodesmium optimize dust utilization by selective collection and retention of nutrient-rich particles
- PMID: 35005537
- PMCID: PMC8718973
- DOI: 10.1016/j.isci.2021.103587
Colonies of the marine cyanobacterium Trichodesmium optimize dust utilization by selective collection and retention of nutrient-rich particles
Abstract
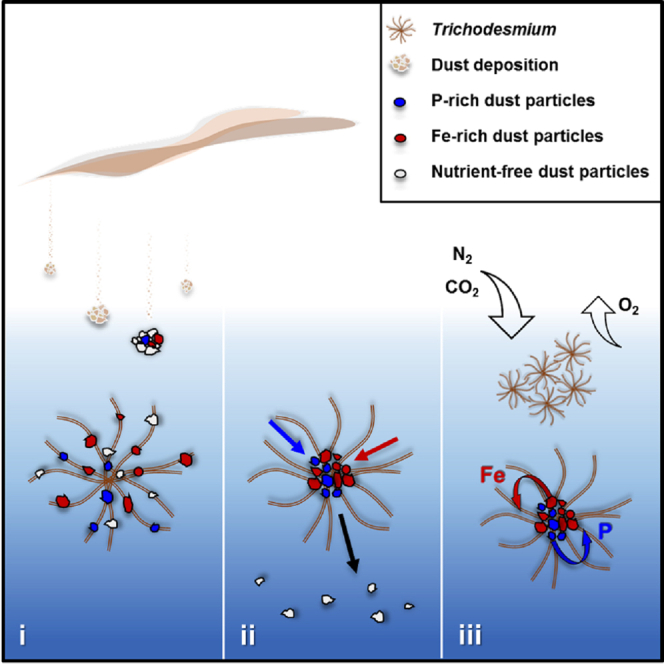
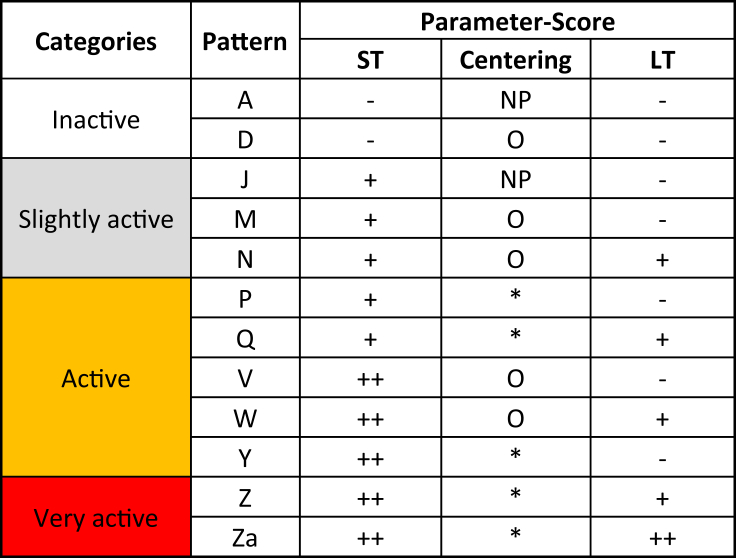
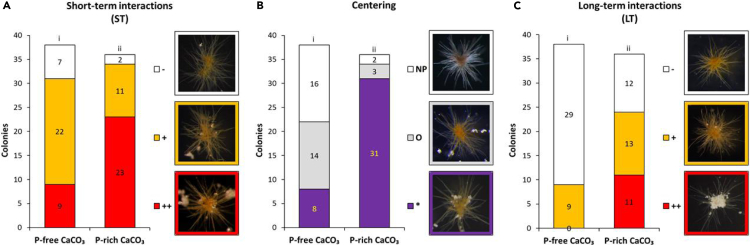
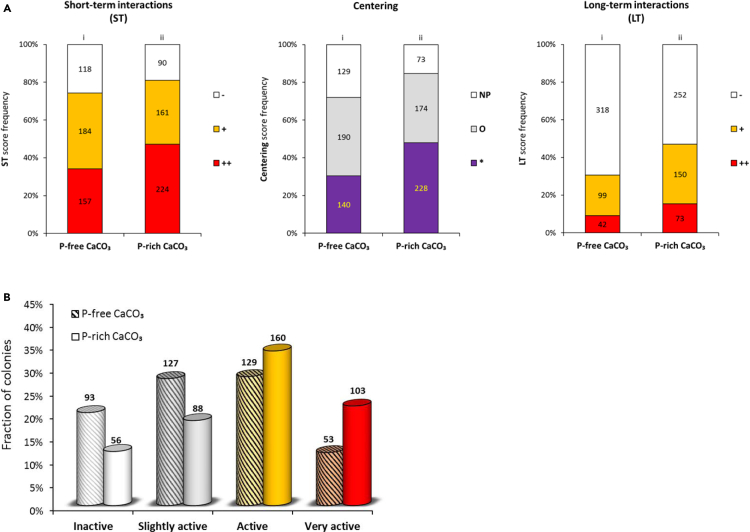
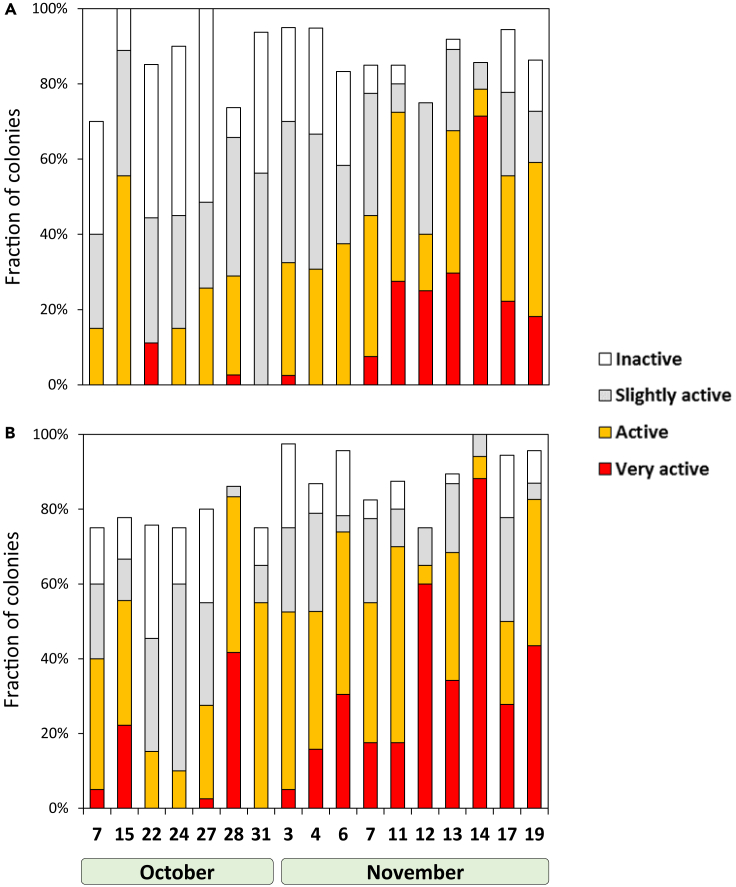
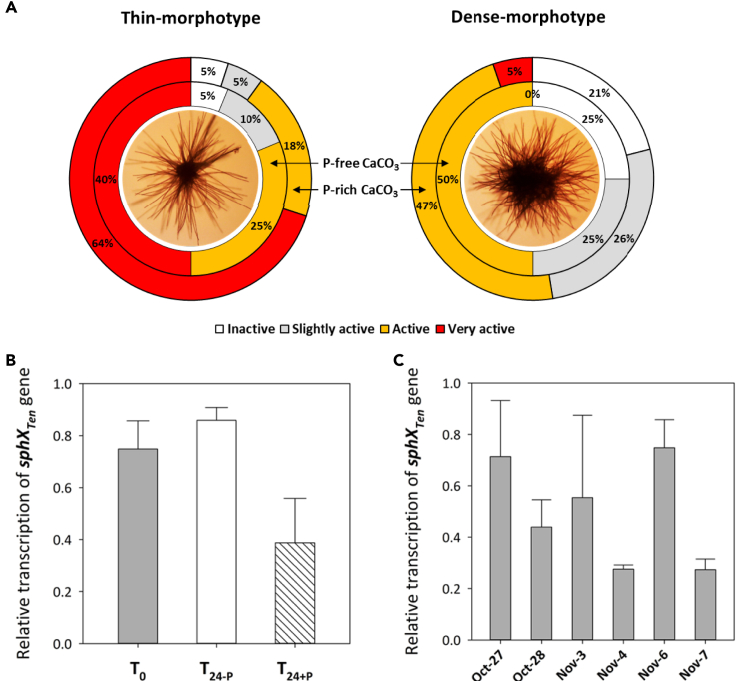
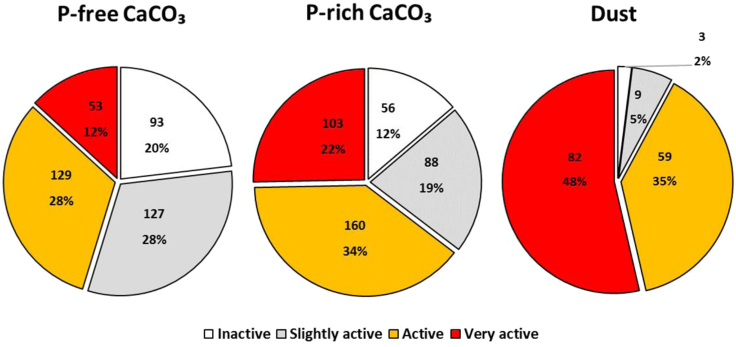
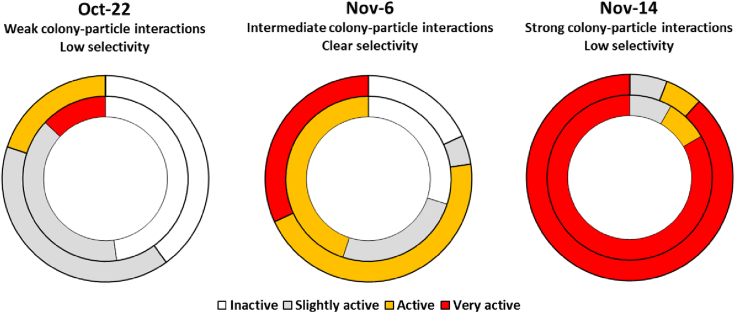
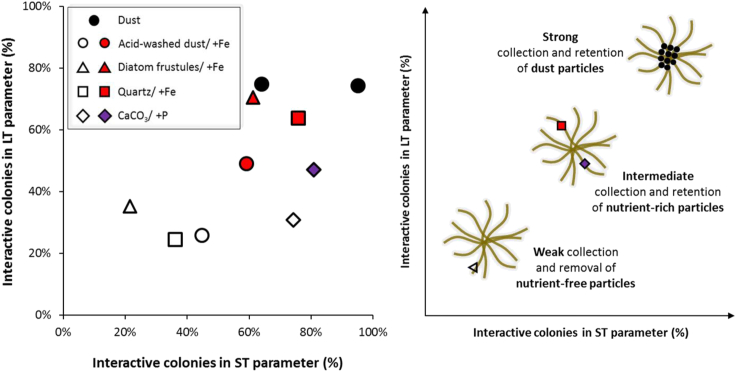
Trichodesmium, a globally important, N2-fixing, and colony-forming cyanobacterium, employs multiple pathways for acquiring nutrients from air-borne dust, including active dust collection. Once concentrated within the colony core, dust can supply Trichodesmium with nutrients. Recently, we reported a selectivity in particle collection enabling Trichodesmium to center iron-rich minerals and optimize its nutrient utilization. In this follow-up study we examined if colonies select Phosphorus (P) minerals. We incubated 1,200 Trichodesmium colonies from the Red Sea with P-free CaCO3, P-coated CaCO3, and dust, over an entire bloom season. These colonies preferably interacted, centered, and retained P-coated CaCO3 compared with P-free CaCO3. In both studies, Trichodesmium clearly favored dust over all other particles tested, whereas nutrient-free particles were barely collected or retained, indicating that the colonies sense the particle composition and preferably collect nutrient-rich particles. This unique ability contributes to Trichodesmium's current ecological success and may assist it to flourish in future warmer oceans.
Keywords: Ecology; Geomicrobiology; Microbiology.
© 2021 The Authors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
-
- Aguilar-Islas A.M., Wu J., Rember R., Johansen A.M., Shank L.M. Dissolution of aerosol-derived iron in seawater: Leach solution chemistry, aerosol type, and colloidal iron fraction. Mar. Chem. 2010;120:25–33. doi: 10.1016/j.marchem.2009.01.011. - DOI
-
- Ammerman J.W., Hood R.R., Case D.A., Cotner J.B. Phosphorus deficiency in the Atlantic: An emerging paradigm in oceanography. Eos, Trans. Am. Geophys. Union. 2003;84:165–170. doi: 10.1029/2003EO180001. - DOI
-
- Anderson L.D., Faul K.L., Paytan A. Phosphorus associations in aerosols: What can they tell us about P bioavailability? Mar. Chem. 2010;120:44–56. doi: 10.1016/j.marchem.2009.04.008. - DOI
-
- Barkley A.E., Prospero J.M., Mahowald N., Hamilton D.S., Popendorf K.J., Oehlert A.M., Pourmand A., Gatineau A., Panechou-Pulcherie K., Blackwelder P., Gaston C.J. African biomass burning is a substantial source of phosphorus deposition to the Amazon, Tropical Atlantic Ocean, and Southern Ocean. Proc. Natl. Acad. Sci. U. S. A. 2019;116:16216–16221. doi: 10.1073/pnas.1906091116. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
