

IL-27 induces IFN/STAT1-dependent genes and enhances function of TIGIT+ HIVGag-specific T cells
- PMID: 35005538
- PMCID: PMC8717455
- DOI: 10.1016/j.isci.2021.103588
IL-27 induces IFN/STAT1-dependent genes and enhances function of TIGIT+ HIVGag-specific T cells
Abstract
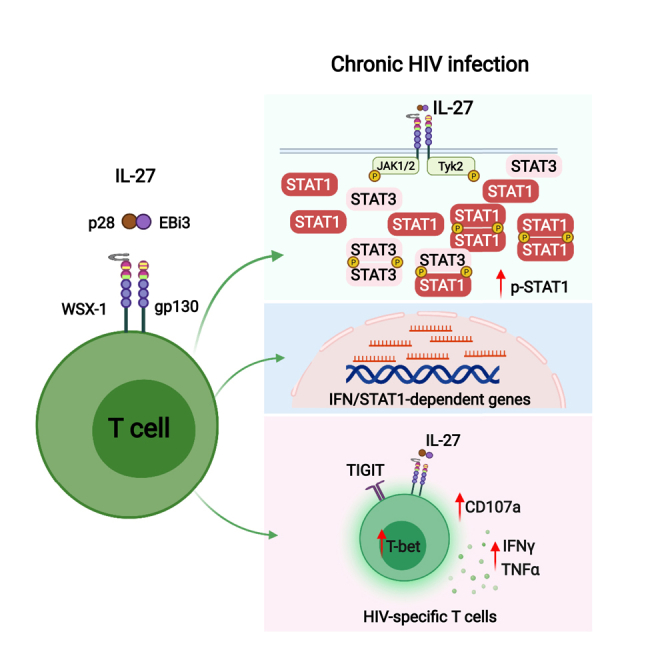
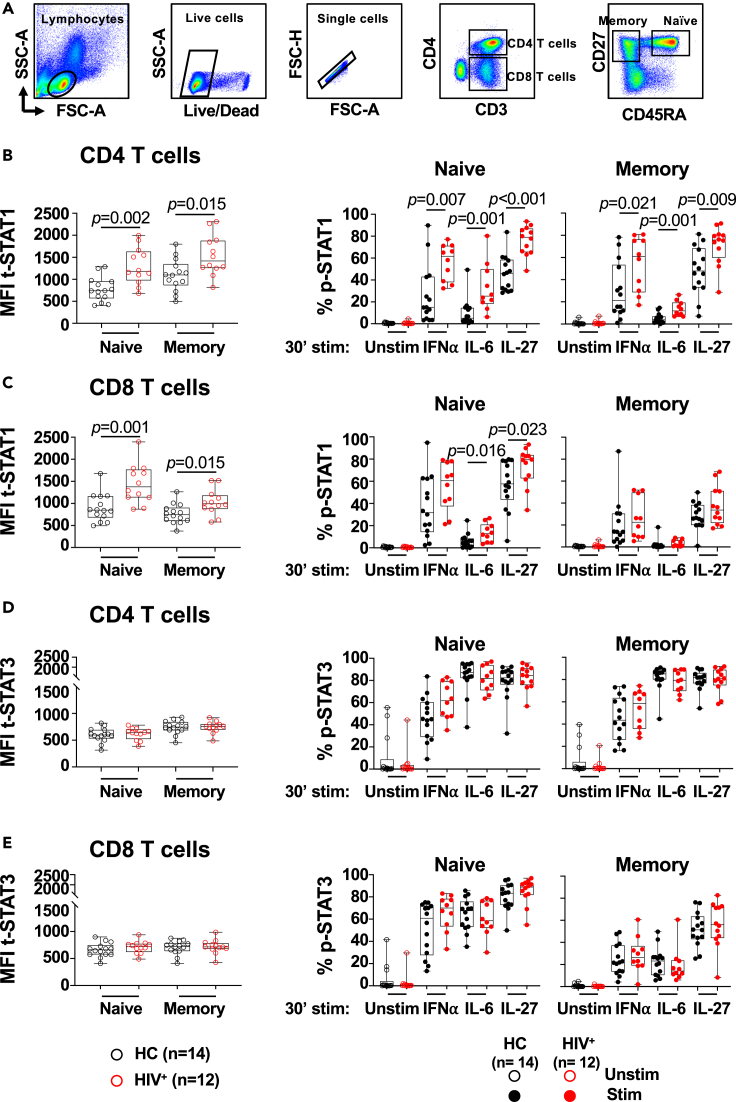
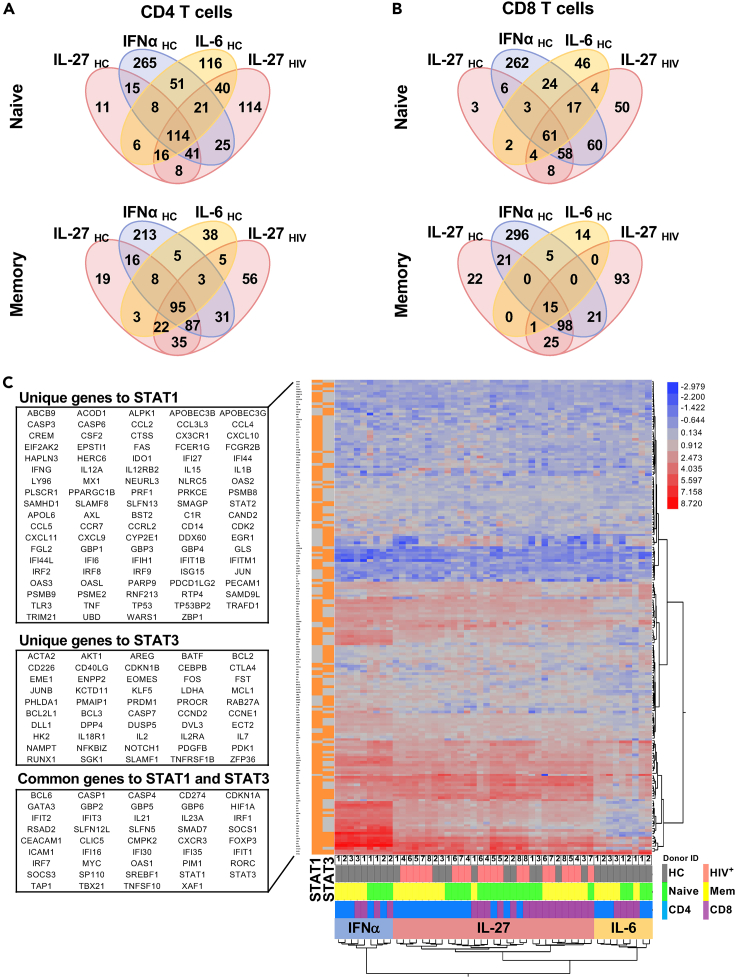
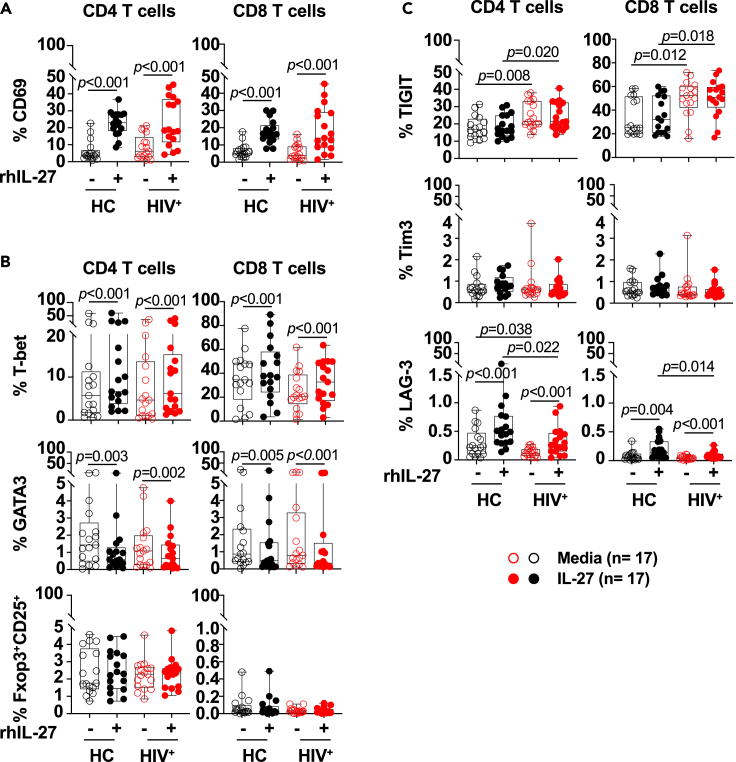
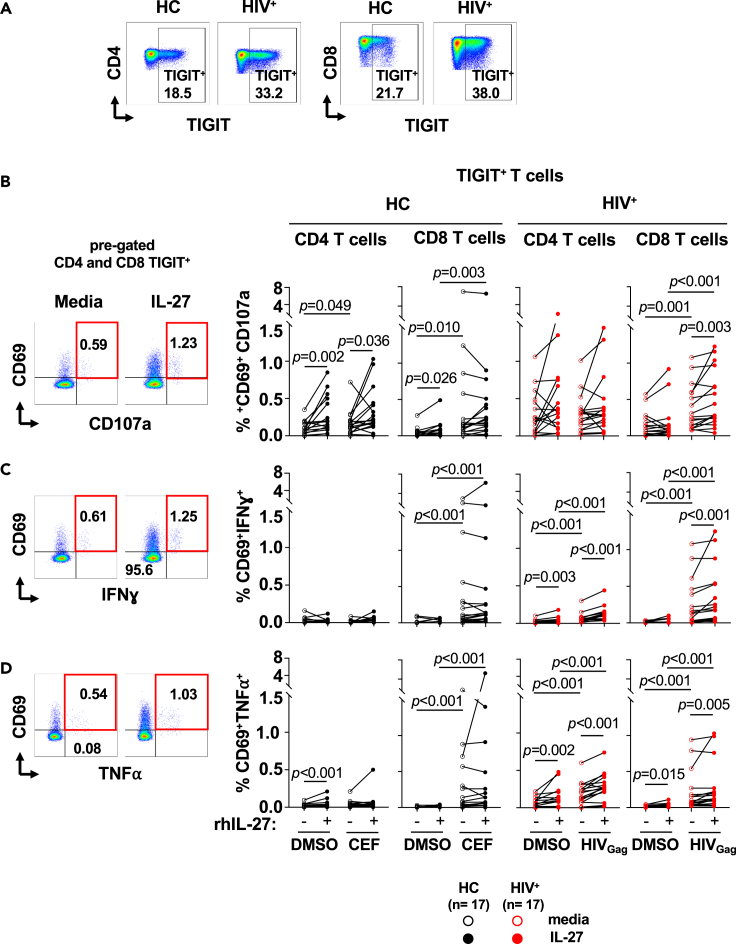
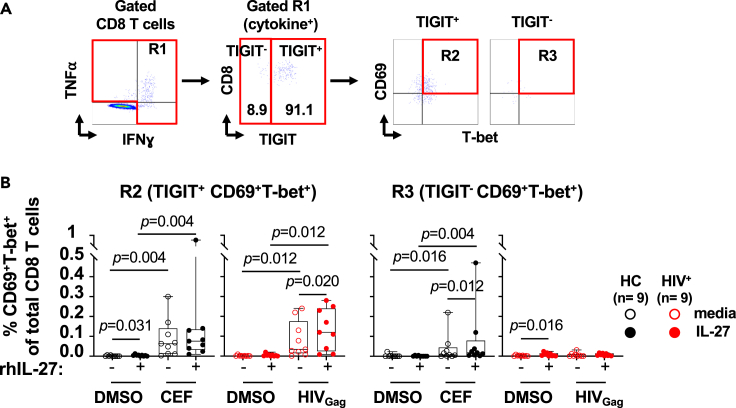
HIV-specific T cells have diminished effector function and fail to control/eliminate the virus. IL-27, a member of the IL-6/IL-12 cytokine superfamily has been shown to inhibit HIV replication. However, whether or not IL-27 can enhance HIV-specific T cell function is largely unknown. In the present manuscript, we investigated the role of IL-27 signaling in human T cells by evaluating the global transcriptional changes related to the function of HIV-specific T cells. We found that T cells from people living with HIV (PLWH), expressed higher levels of STAT1 leading to enhanced STAT1 activation upon IL-27 stimulation. Observed IL-27 induced transcriptional changes were associated with IFN/STAT1-dependent pathways in CD4 and CD8 T cells. Importantly, IL-27 dependent modulation of T-bet expression promoted IFNγ secretion by TIGIT+HIVGag-specific T cells. This new immunomodulatory effect of IL-27 on HIV-specific T cell function suggests its potential therapeutic use in cure strategies.
Keywords: Components of the immune system; Immune response; Transcriptomics.
© 2021.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Batten M., Kljavin N.M., Li J., Walter M.J., de Sauvage F.J., Ghilardi N. Cutting edge: IL-27 is a potent inducer of IL-10 but not FoxP3 in murine T cells. J. Immunol. 2008;180:2752–2756. - PubMed
-
- Buggert M., Nguyen S., McLane L.M., Steblyanko M., Anikeeva N., Paquin-Proulx D., Del Rio Estrada P.M., Ablanedo-Terrazas Y., Noyan K., Reuter M.A., et al. Limited immune surveillance in lymphoid tissue by cytolytic CD4+ T cells during health and HIV disease. PLoS Pathog. 2018;14:e1006973. - PMC - PubMed
-
- Catalfamo M., Di Mascio M., Hu Z., Srinivasula S., Thaker V., Adelsberger J., Rupert A., Baseler M., Tagaya Y., Roby G., et al. HIV infection-associated immune activation occurs by two distinct pathways that differentially affect CD4 and CD8 T cells. Proc. Natl. Acad. Sci. U S A. 2008;105:19851–19856. - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
