

Systematic Genetic Interaction Analysis Identifies a Transcription Factor Circuit Required for Oropharyngeal Candidiasis
- PMID: 35012341
- PMCID: PMC8749425
- DOI: 10.1128/mbio.03447-21
Systematic Genetic Interaction Analysis Identifies a Transcription Factor Circuit Required for Oropharyngeal Candidiasis
Abstract
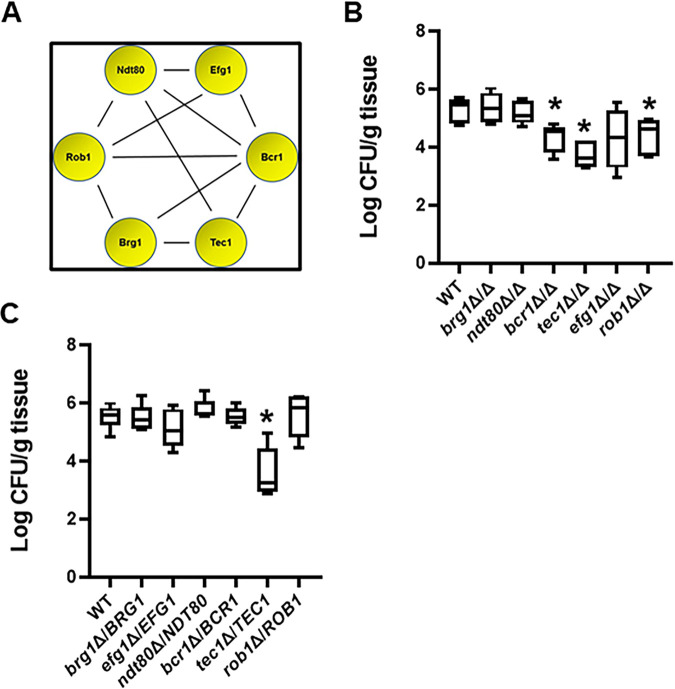
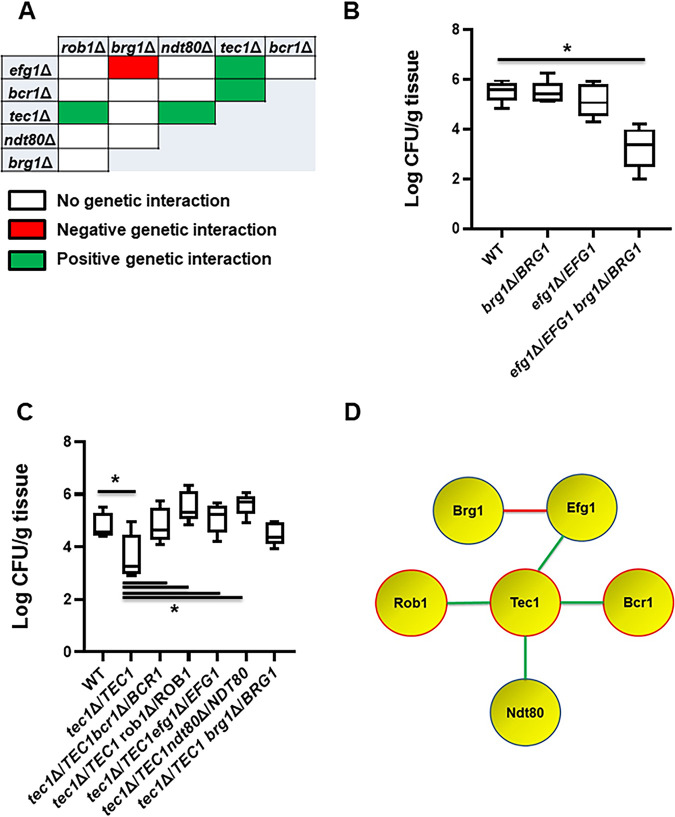
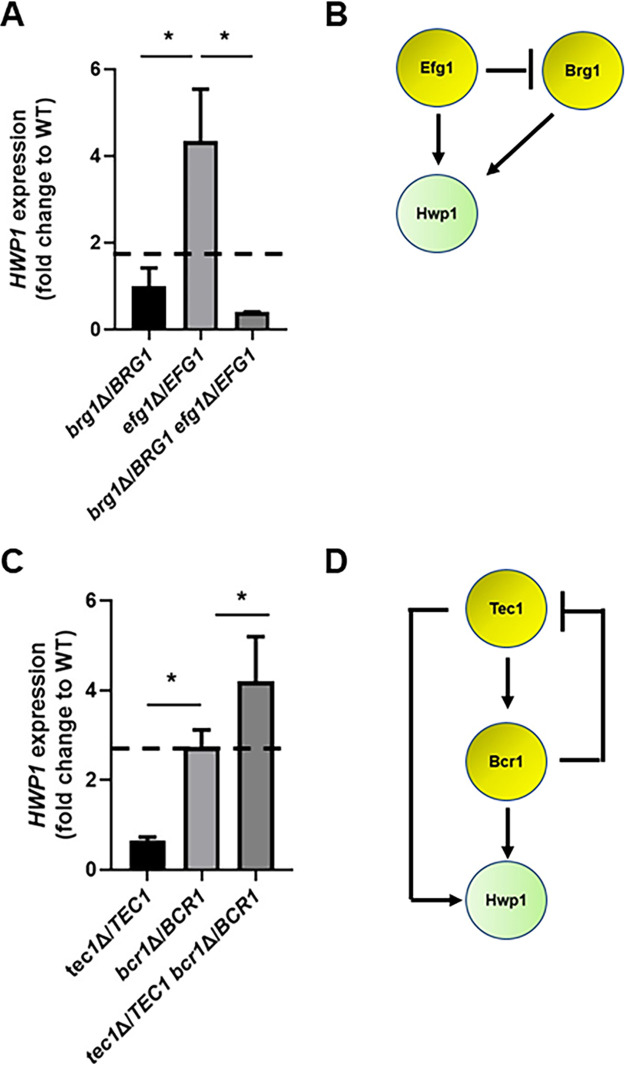
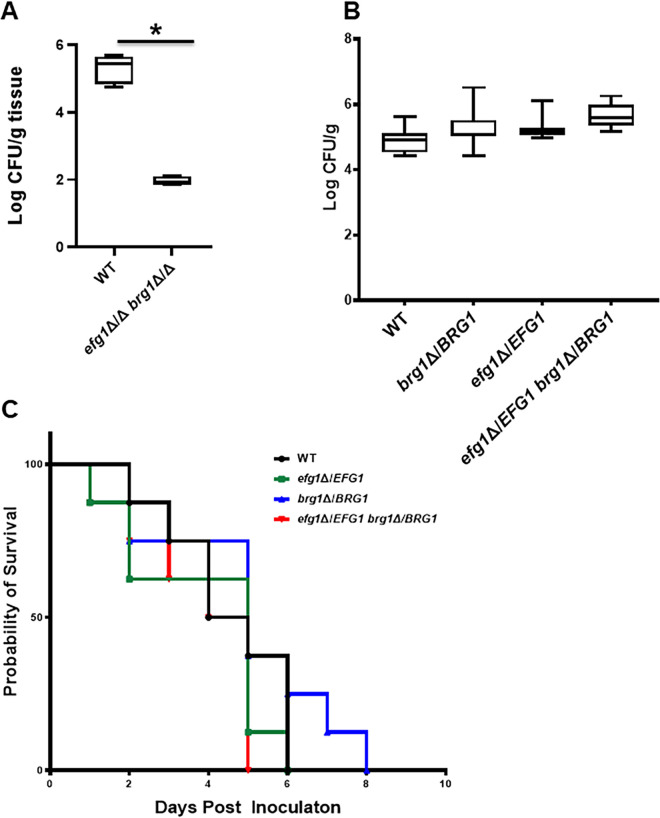
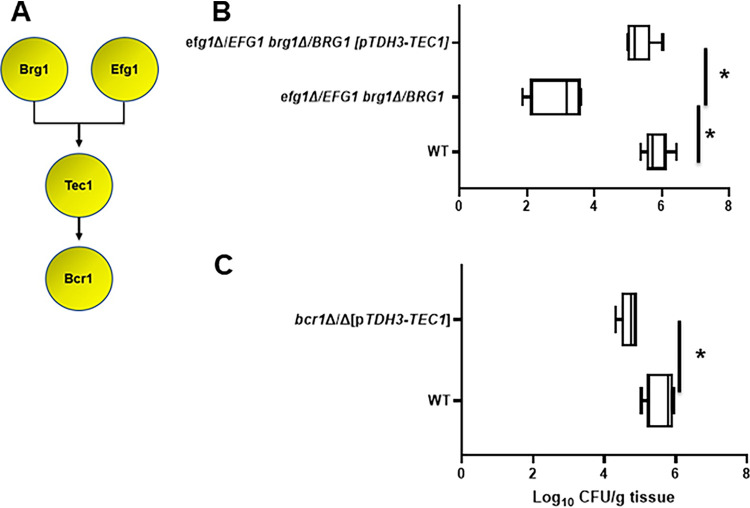
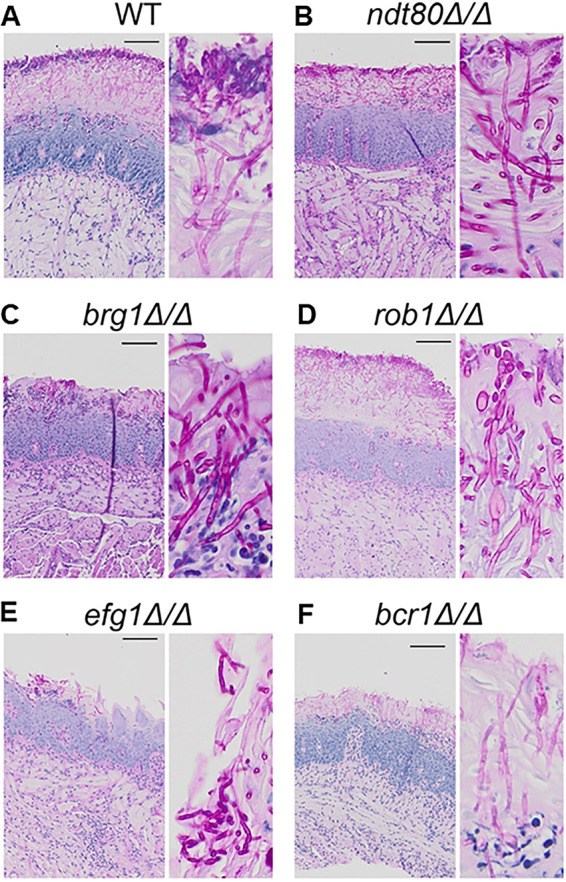
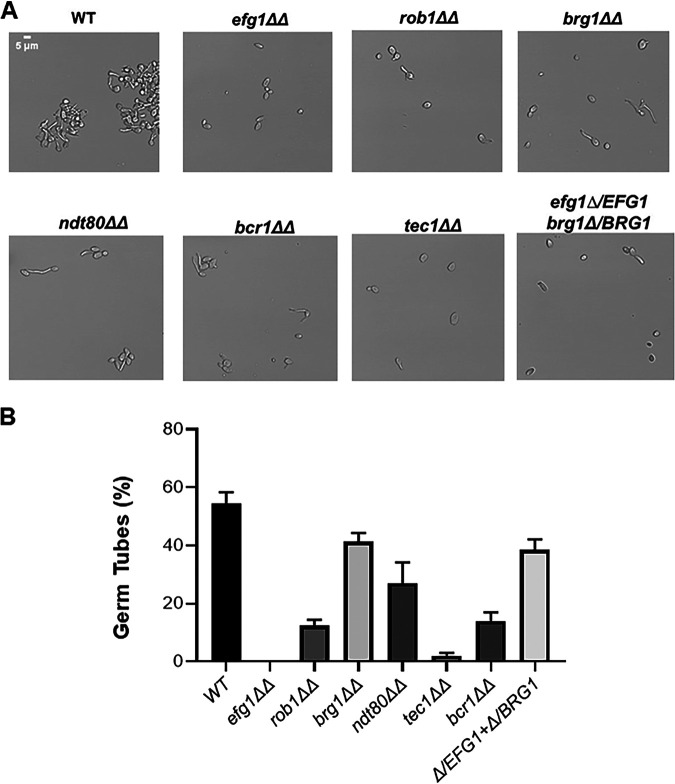
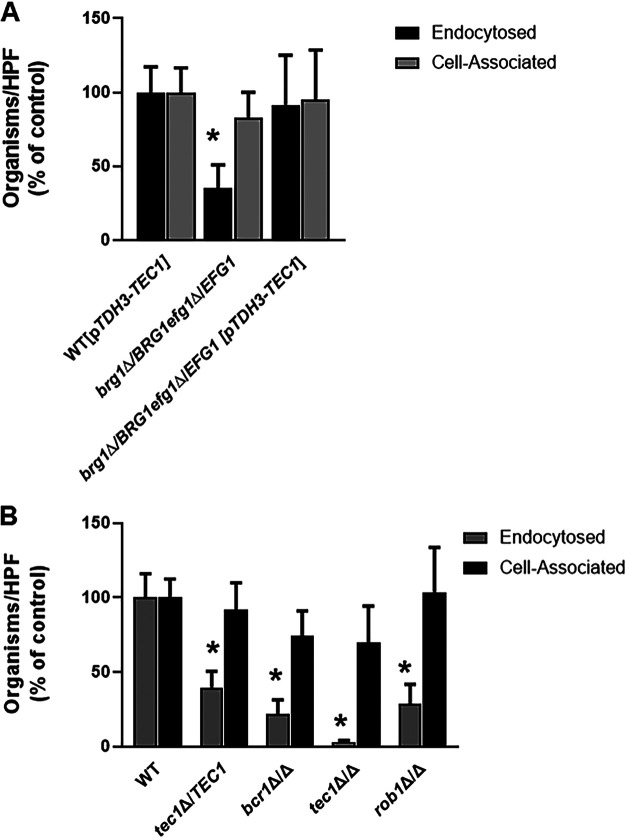
Oropharyngeal candidiasis (OPC) is a common infection that complicates a wide range of medical conditions and can cause either mild or severe disease depending on the patient. The pathobiology of OPC shares many features with candidal biofilms of abiotic surfaces. The transcriptional regulation of C. albicans biofilm formation on abiotic surfaces has been extensively characterized and involves six key transcription factors (Efg1, Ndt80, Rob1, Bcr1, Brg1, and Tec1). To determine if the in vitro biofilm transcriptional regulatory network also plays a role in OPC, we carried out a systematic genetic interaction analysis in a mouse model of C. albicans OPC. Whereas each of the six transcription factors are required for in vitro biofilm formation, only three homozygous deletion mutants (tec1ΔΔ, bcr1ΔΔ, and rob1ΔΔ) and one heterozygous mutant (tec1Δ/TEC1) have reduced infectivity in the mouse model of OPC. Although single mutants (heterozygous or homozygous) of BRG1 and EFG1 have no effect on fungal burden, double heterozygous and homozygous mutants have dramatically reduced infectivity, indicating a critical genetic interaction between these two transcription factors during OPC. Using epistasis analysis, we have formulated a genetic circuit, [EFG1+BRG1]→TEC1→BCR1, that is required for OPC infectivity and oral epithelial cell endocytosis. Surprisingly, we also found transcription factor mutants with in vitro defects in filamentation, such as efg1ΔΔ, rob1ΔΔ, and brg1ΔΔ filament, during oral infection and that reduced filamentation does not correlate with infectivity. Taken together, these data indicate that key in vitro biofilm transcription factors are involved in OPC but that the network characteristics and functional connections during infection are distinct from those observed in vivo. IMPORTANCE The pathology of oral candidiasis has features of biofilm formation, a well-studied process in vitro. Based on that analogy, we hypothesized that the network of transcription factors that regulates in vitro biofilm formation has similarities and differences during oral infection. To test this, we employed the first systematic genetic interaction analysis of C. albicans in a mouse model of oropharyngeal infection. This revealed that the six regulators involved in in vitro biofilm formation played roles in vivo but that the functional connections between factors were quite distinct. Surprisingly, we also found that while many of the factors are required for filamentation in vitro, none of the transcription factor deletion mutants was deficient for this key virulence trait in vivo. These observations clearly demonstrate that C. albicans regulates key aspects of its biology differently in vitro and in vivo.
Keywords: Candida albicans; biofilms; complex haploinsufficiency; oropharyngeal candidiasis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Break TJ, Oikonomou V, Dutzan N, Desai JV, Swidergall M, Freiwald T, Chauss D, Harrison OJ, Alejo J, Williams DW, Pittaluga S, Lee CR, Bouladoux N, Swamydas M, Hoffman KW, Greenwell-Wild T, Bruno VM, Rosen LB, Lwin W, Renteria A, Pontejo SM, Shannon JP, Myles IA, Olbrich P, Ferré EMN, Schmitt M, Martin D, Barber DL, Solis NV, Notarangelo LD, Serreze DV, Matsumoto M, Hickman HD, Murphy PM, Anderson MS, Lim JK, Holland SM, Filler SG, Afzali B, Belkaid Y, Moutsopoulos NM, Lionakis MS, Genomics and Computational Biology Core . 2021. Aberrant type I immunity drives susceptibility to mucosal fungal infections. Science 371:5731. doi: 10.1126/science.aay5731. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
