

Migraine-relevant sex-dependent activation of mouse meningeal afferents by TRPM3 agonists
- PMID: 35012445
- PMCID: PMC8903645
- DOI: 10.1186/s10194-021-01383-8
Migraine-relevant sex-dependent activation of mouse meningeal afferents by TRPM3 agonists
Erratum in
-
Correction to: Migraine-relevant sex-dependent activation of mouse meningeal afferents by TRPM3 agonists.J Headache Pain. 2022 Jan 28;23(1):17. doi: 10.1186/s10194-022-01390-3. J Headache Pain. 2022. PMID: 35090400 Free PMC article. No abstract available.
Abstract
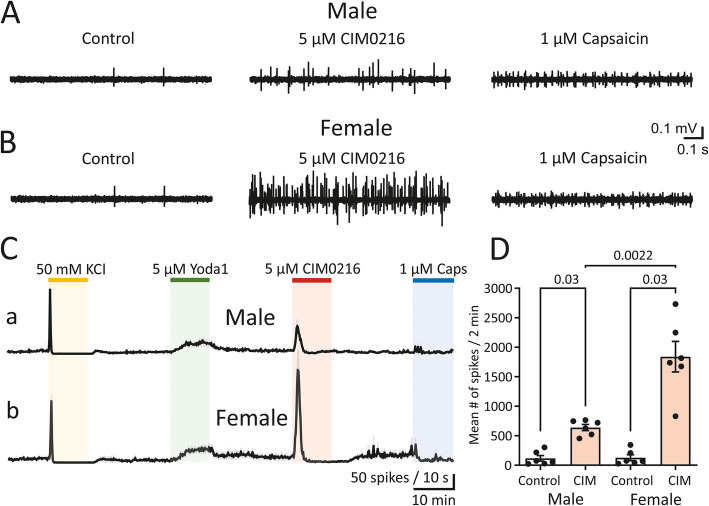
Background: Migraine is a common brain disorder that predominantly affects women. Migraine pain seems mediated by the activation of mechanosensitive channels in meningeal afferents. Given the role of transient receptor potential melastatin 3 (TRPM3) channels in mechanical activation, as well as hormonal regulation, these channels may play a role in the sex difference in migraine. Therefore, we investigated whether nociceptive firing induced by TRPM3 channel agonists in meningeal afferents was different between male and female mice. In addition, we assessed the relative contribution of mechanosensitive TRPM3 channels and that of mechanosensitive Piezo1 channels and transient receptor potential vanilloid 1 (TRPV1) channels to nociceptive firing relevant to migraine in both sexes.
Methods: Ten- to 13-week-old male and female wildtype (WT) C57BL/6 J mice were used. Nociceptive spikes were recorded directly from nerve terminals in the meninges in the hemiskull preparations.
Results: Selective agonists of TRPM3 channels profoundly activated peripheral trigeminal nerve fibres in mouse meninges. A sex difference was observed for nociceptive firing induced by either PregS or CIM0216, both agonists of TRPM3 channels, with the induced firing being particularly prominent for female mice. Application of Yoda1, an agonist of Piezo1 channels, or capsaicin activating TRPV1 channels, although also leading to increased nociceptive firing of meningeal fibres, did not reveal a sex difference. Cluster analyses of spike activities indicated a massive and long-lasting activation of TRPM3 channels with preferential induction of large-amplitude spikes in female mice. Additional spectral analysis revealed a dominant contribution of spiking activity in the α- and β-ranges following TRPM3 agonists in female mice.
Conclusions: Together, we revealed a specific mechanosensitive profile of nociceptive firing in females and suggest TRPM3 channels as a potential novel candidate for the generation of migraine pain, with particular relevance to females.
Keywords: CIM0216; Migraine; Nociception; Pregnenolone sulfate; Sex-dependence; TRPM3.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





References
-
- Global Burden of Disease Study (2019) US Seattle, Institute for Health Metrics and Evaluation http://ghdxhealthdataorg/gbd-results-tool Accessed 15 Oct 2020
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
