

Chromatin basis of the senescence-associated secretory phenotype
- PMID: 35012849
- PMCID: PMC9106822
- DOI: 10.1016/j.tcb.2021.12.003
Chromatin basis of the senescence-associated secretory phenotype
Abstract
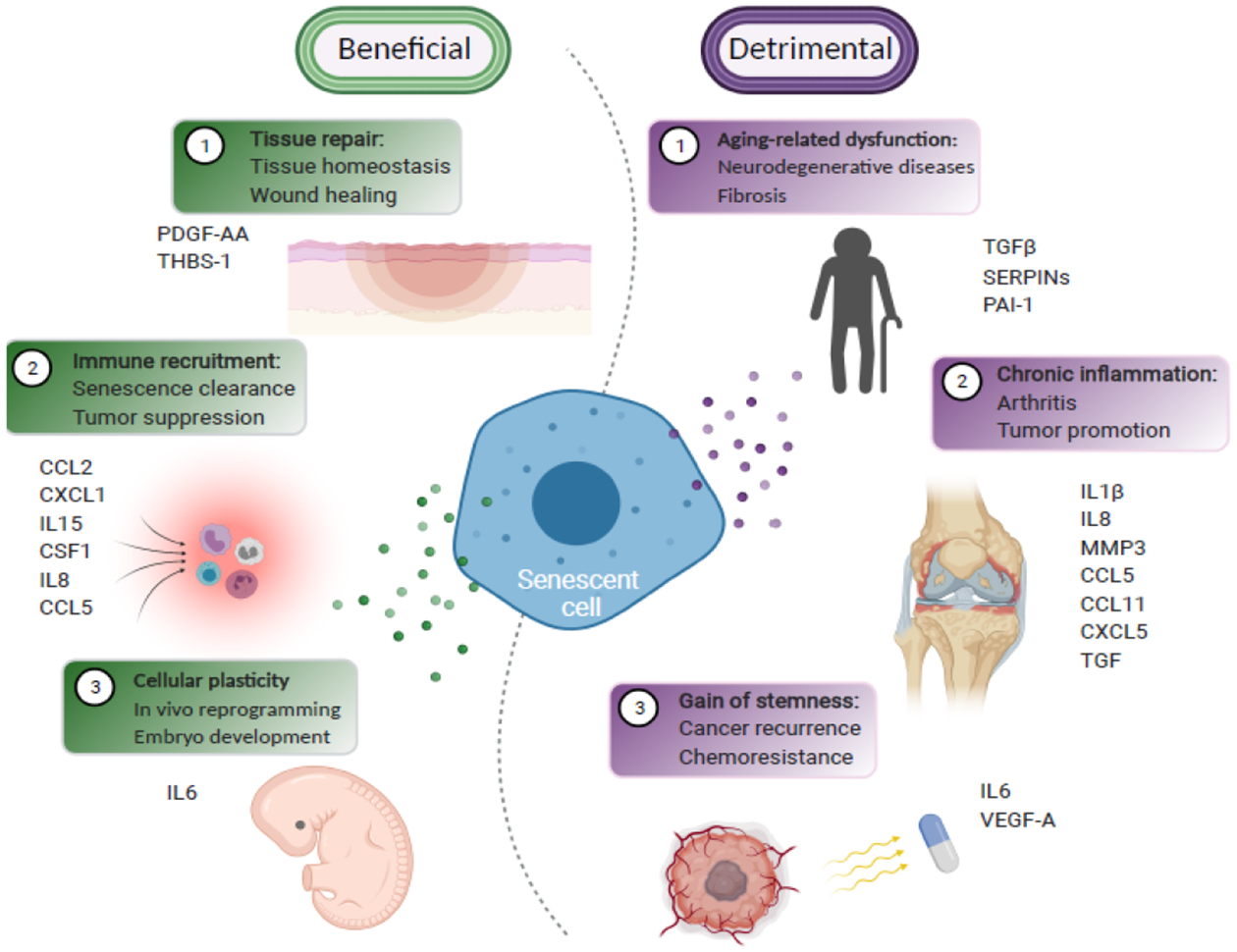
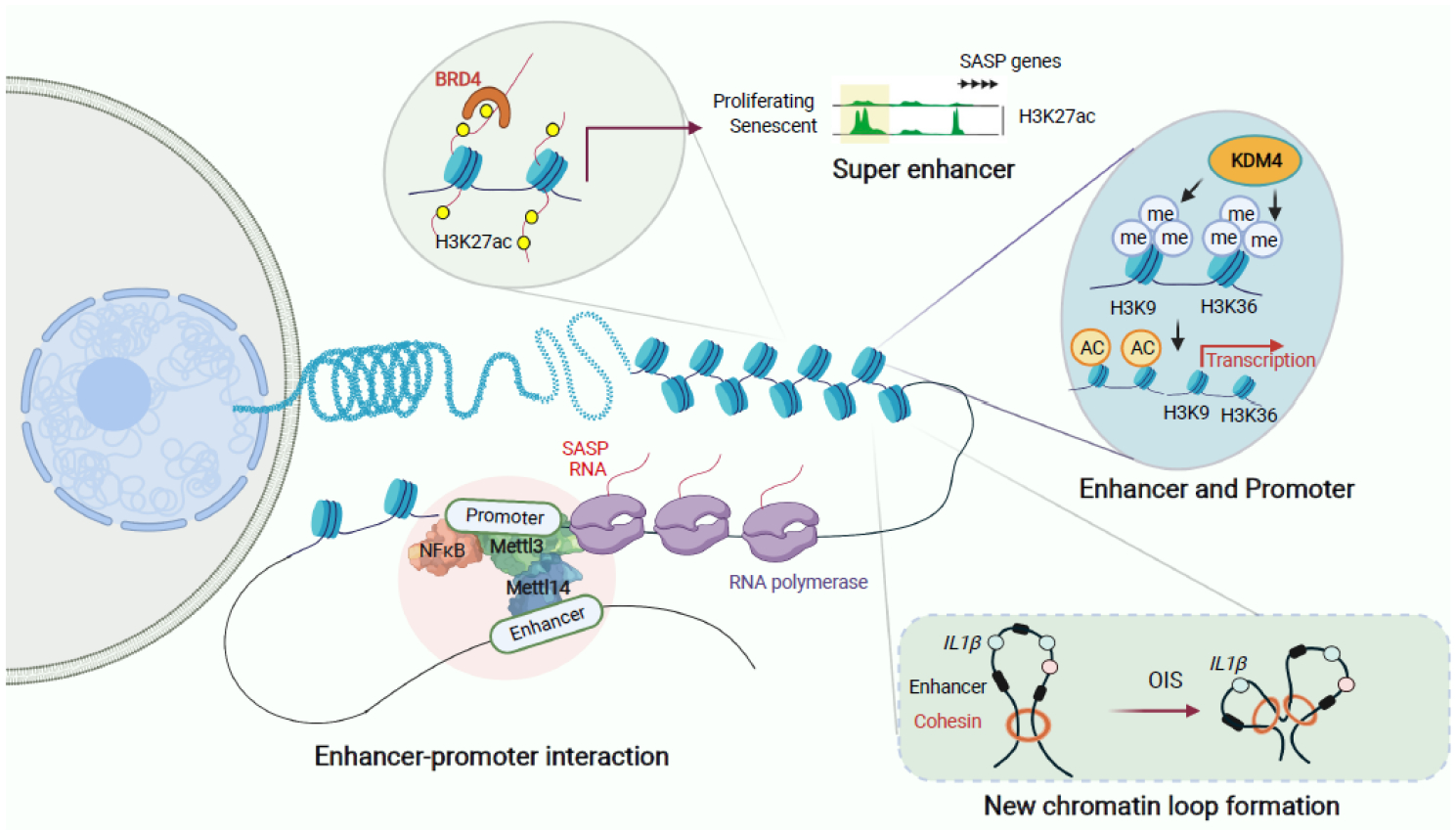
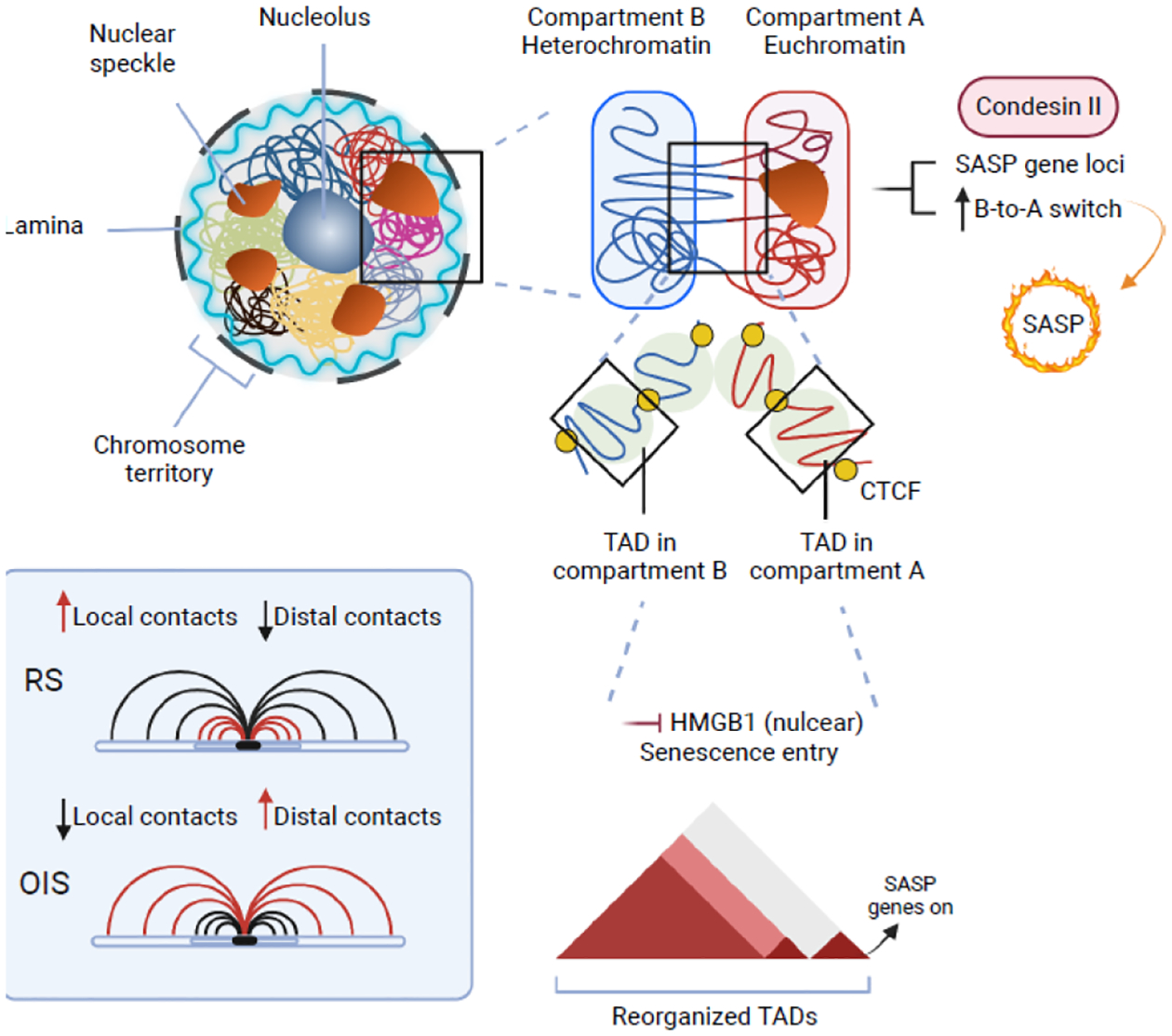
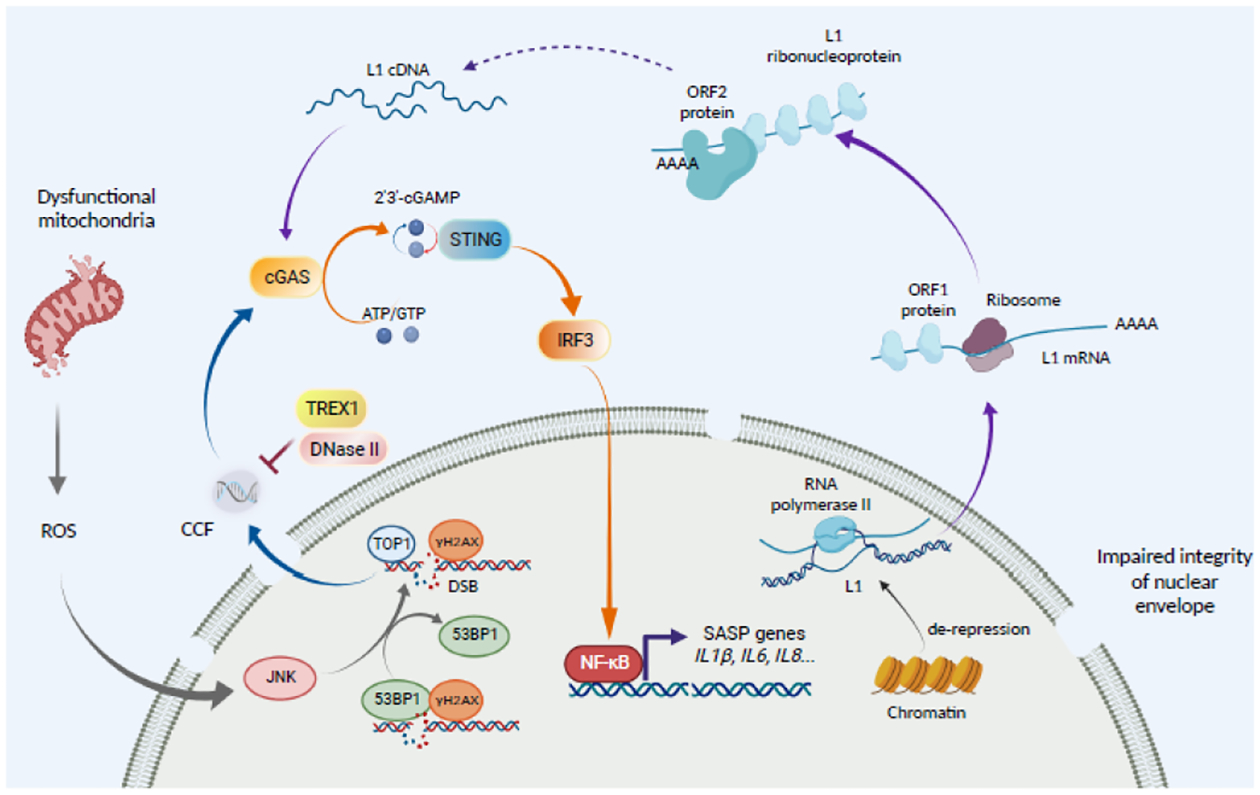
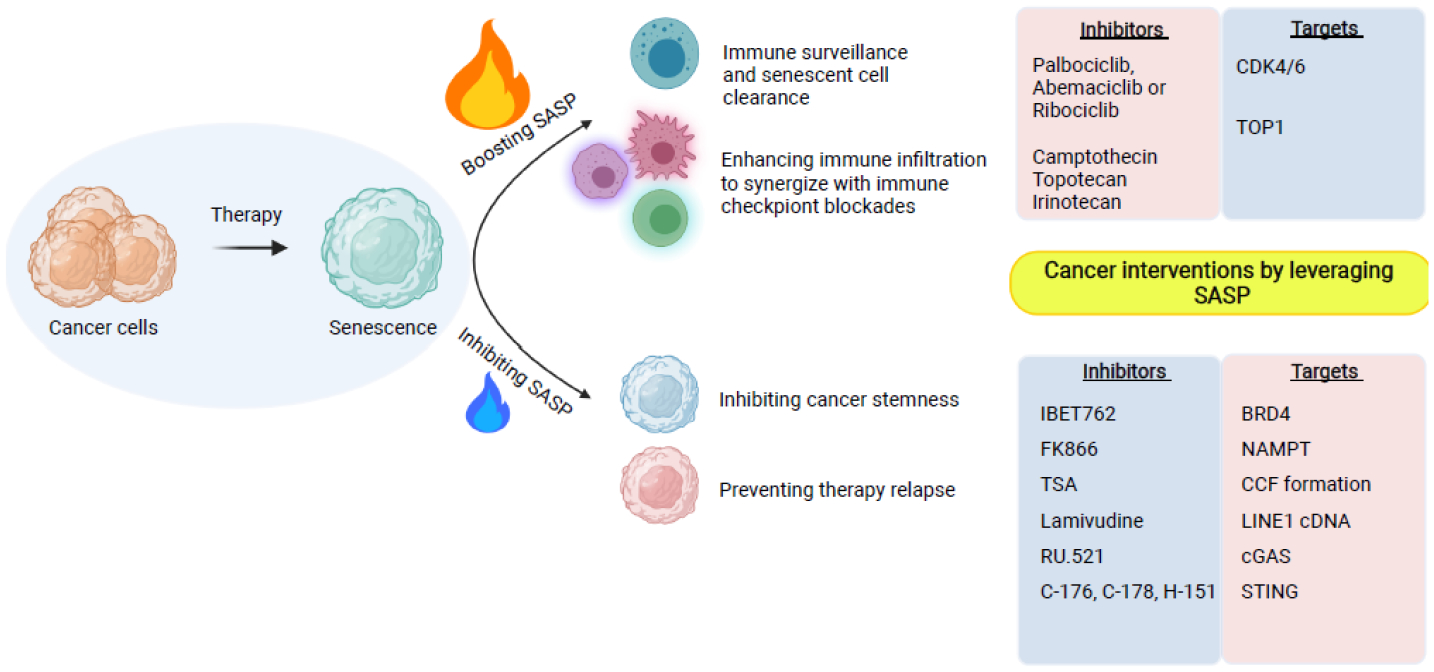
Cellular senescence is a stable cell growth arrest. Senescent cells are metabolically active, as exemplified by the secretion of inflammatory cytokines, chemokines, and growth factors, which is termed senescence-associated secretory phenotype (SASP). The SASP exerts a range of functions in both normal health and pathology, which is possibly best characterized in cancers and physical aging. Recent studies demonstrated that chromatin is instrumental in regulating the SASP both through nuclear transcription and via the innate immune cyclic GMP-AMP synthase (cGAS)-stimulator of interferon genes (STING) pathway in the cytoplasm. Here, we will review these regulatory mechanisms, with an emphasis on most recent developments in the field. We will highlight the challenges and opportunities in developing intervention approaches, such as targeting chromatin regulatory mechanisms, to alter the SASP as an emerging approach to combat cancers and achieve healthy aging.
Keywords: chromatin structure; cytoplasmic chromatin; enhancer–promoter interaction; senescence-associated secretory phenotype; senomorphics.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Hayflick L (1965) The Limited in Vitro Lifetime of Human Diploid Cell Strains. Exp Cell Res 37, 614–36. - PubMed
-
- Kuilman T et al. (2008) Oncogene-induced senescence relayed by an interleukin-dependent inflammatory network. Cell 133 (6), 1019–31. - PubMed
-
- Acosta JC et al. (2008) Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell 133 (6), 1006–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
