

Context-independent encoding of passive and active self-motion in vestibular afferent fibers during locomotion in primates
- PMID: 35013266
- PMCID: PMC8748921
- DOI: 10.1038/s41467-021-27753-z
Context-independent encoding of passive and active self-motion in vestibular afferent fibers during locomotion in primates
Abstract
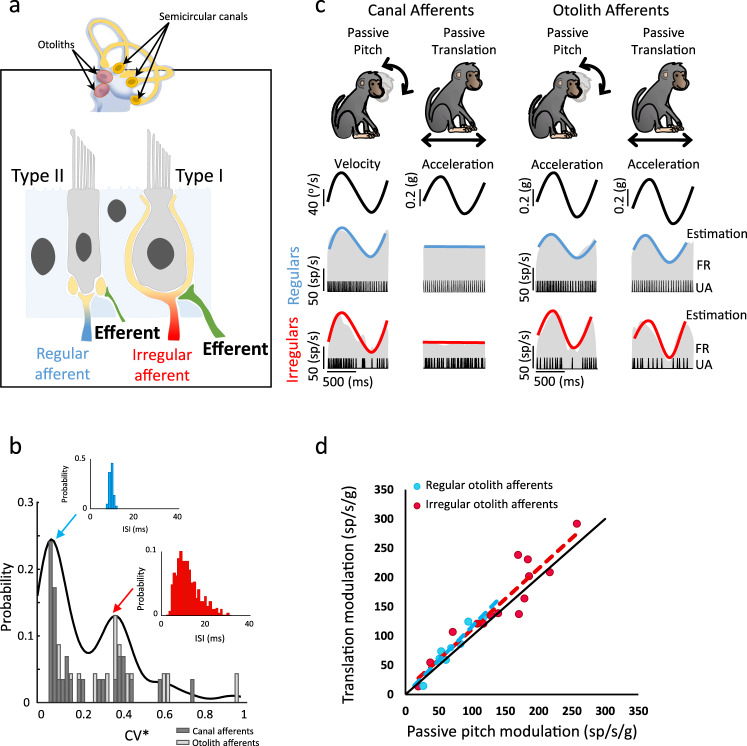
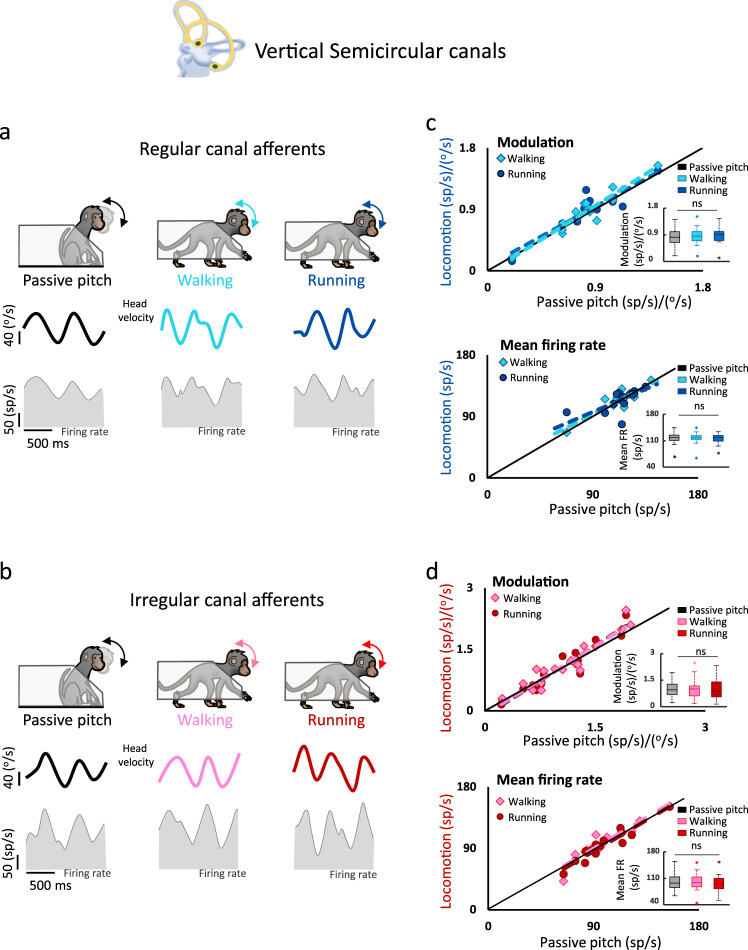
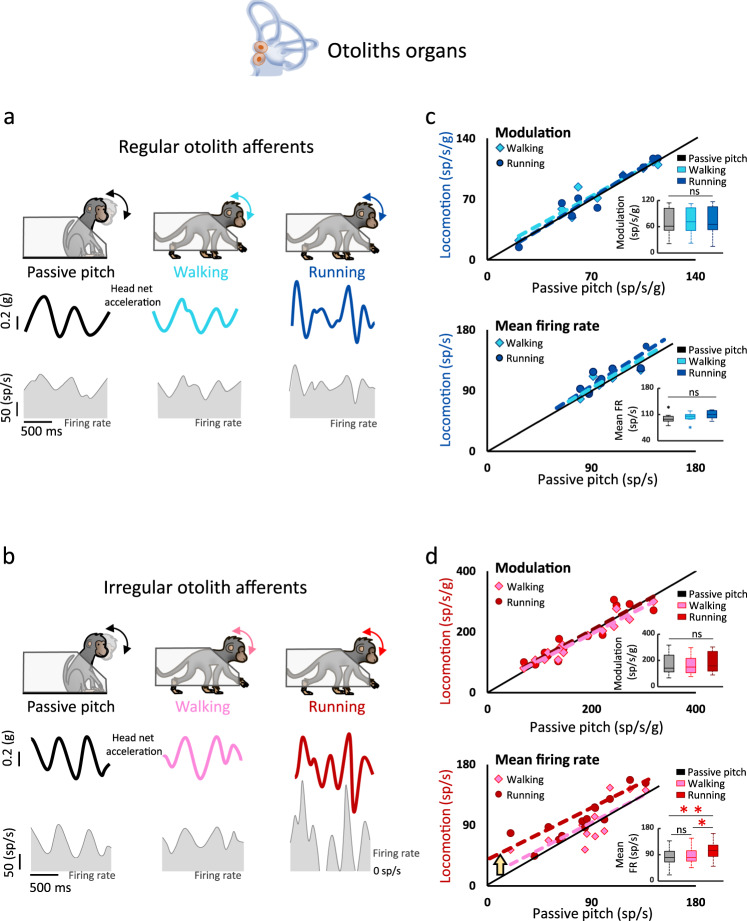
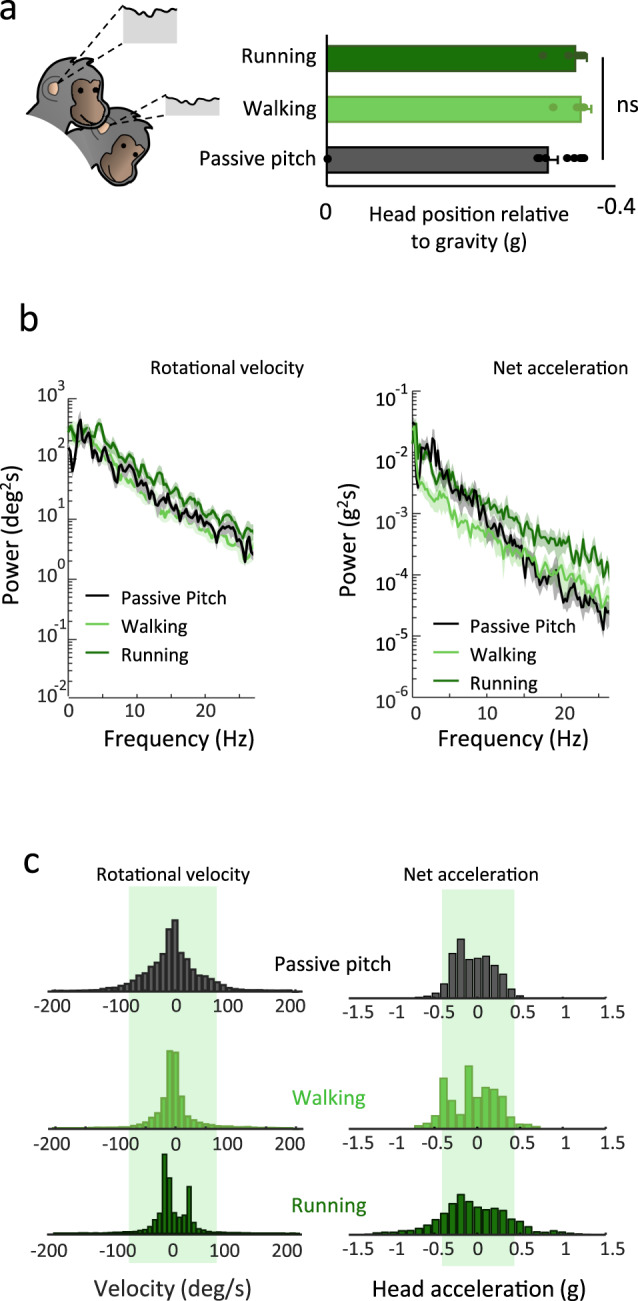
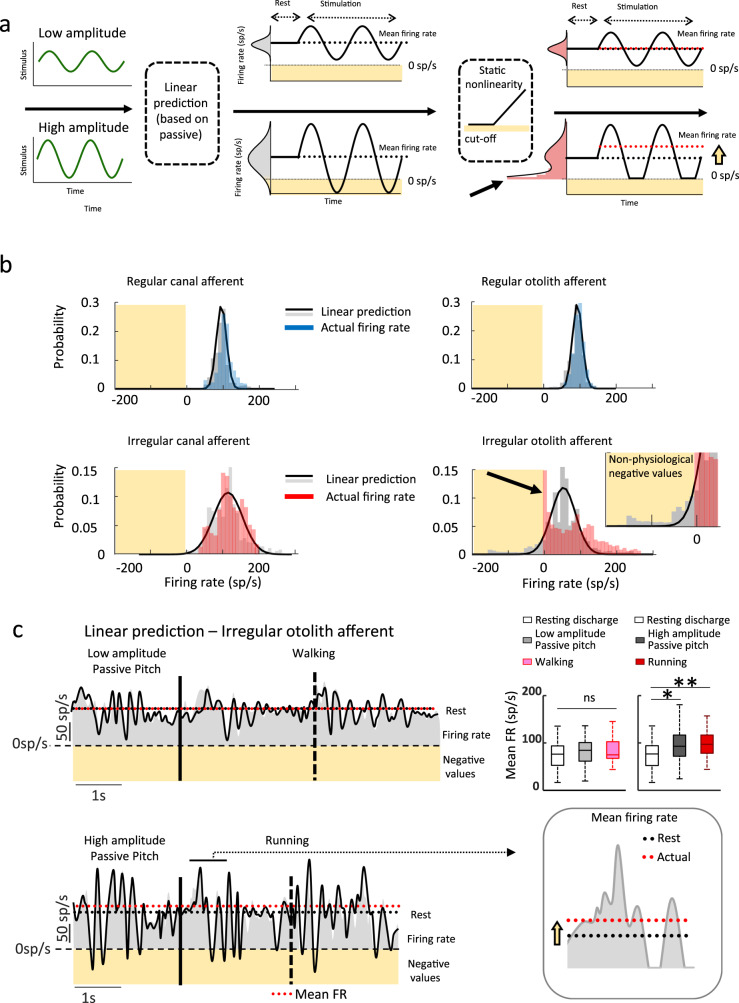
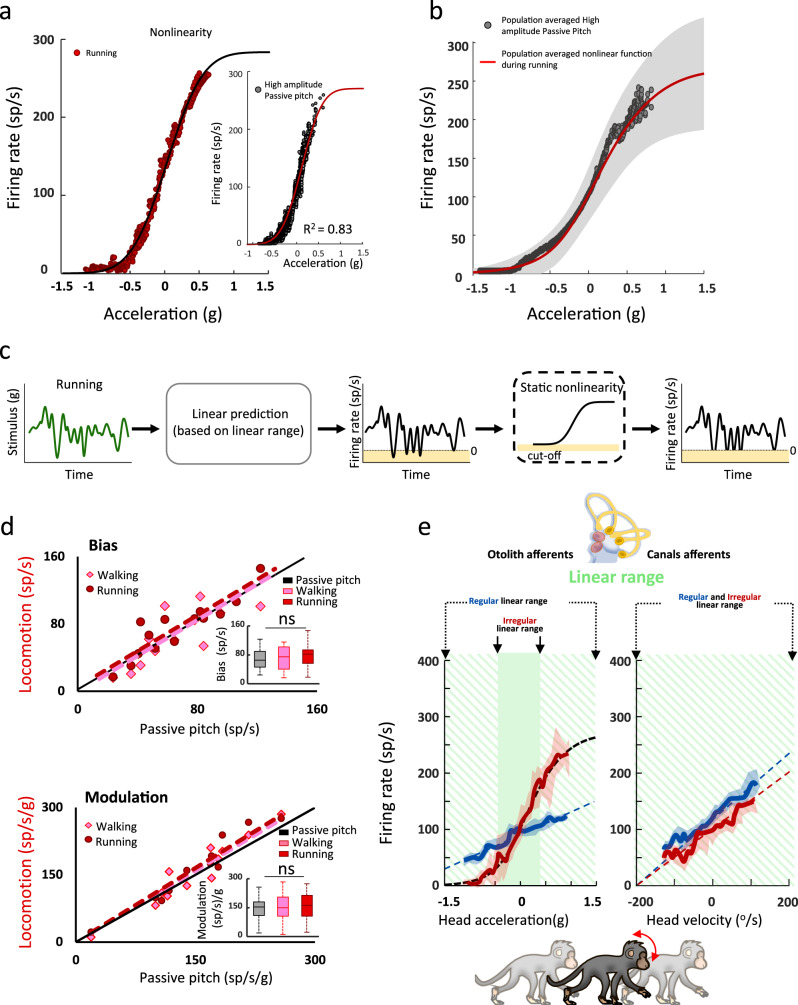
The vestibular system detects head motion to coordinate vital reflexes and provide our sense of balance and spatial orientation. A long-standing hypothesis has been that projections from the central vestibular system back to the vestibular sensory organs (i.e., the efferent vestibular system) mediate adaptive sensory coding during voluntary locomotion. However, direct proof for this idea has been lacking. Here we recorded from individual semicircular canal and otolith afferents during walking and running in monkeys. Using a combination of mathematical modeling and nonlinear analysis, we show that afferent encoding is actually identical across passive and active conditions, irrespective of context. Thus, taken together our results are instead consistent with the view that the vestibular periphery relays robust information to the brain during primate locomotion, suggesting that context-dependent modulation instead occurs centrally to ensure that coding is consistent with behavioral goals during locomotion.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Lysakowski, A. & Goldberg, J. M. In The Vestibular System (eds. Stephen M. Highstein, Richard R. Fay, & Arthur N. Popper) 57–152 (Springer New York, 2004).
-
- Meredith GE. Comparative view of the central organization of afferent and efferent circuitry for the inner ear. Acta Biol. Hung. 1988;39:229–249. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
