

Blockade or Deletion of IFNγ Reduces Macrophage Activation without Compromising CAR T-cell Function in Hematologic Malignancies
- PMID: 35015685
- PMCID: PMC9414118
- DOI: 10.1158/2643-3230.BCD-21-0181
Blockade or Deletion of IFNγ Reduces Macrophage Activation without Compromising CAR T-cell Function in Hematologic Malignancies
Abstract
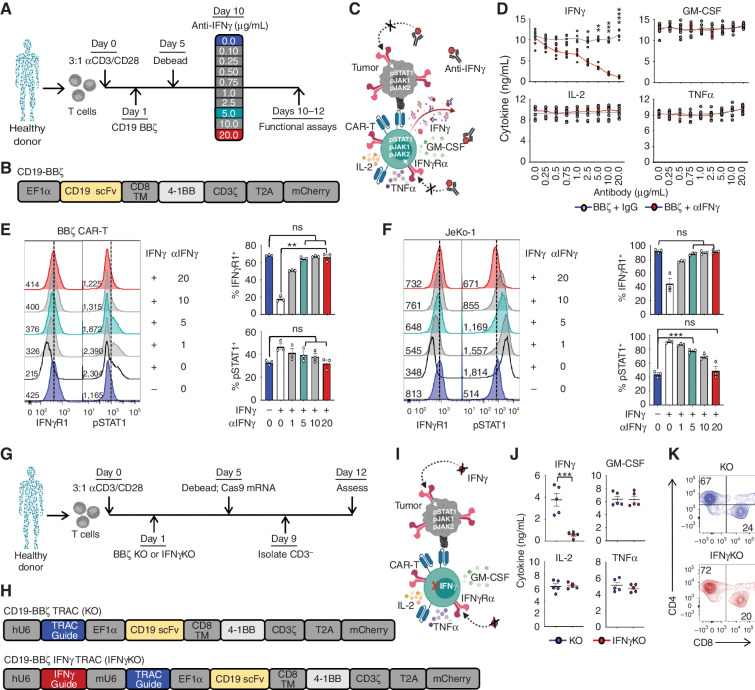
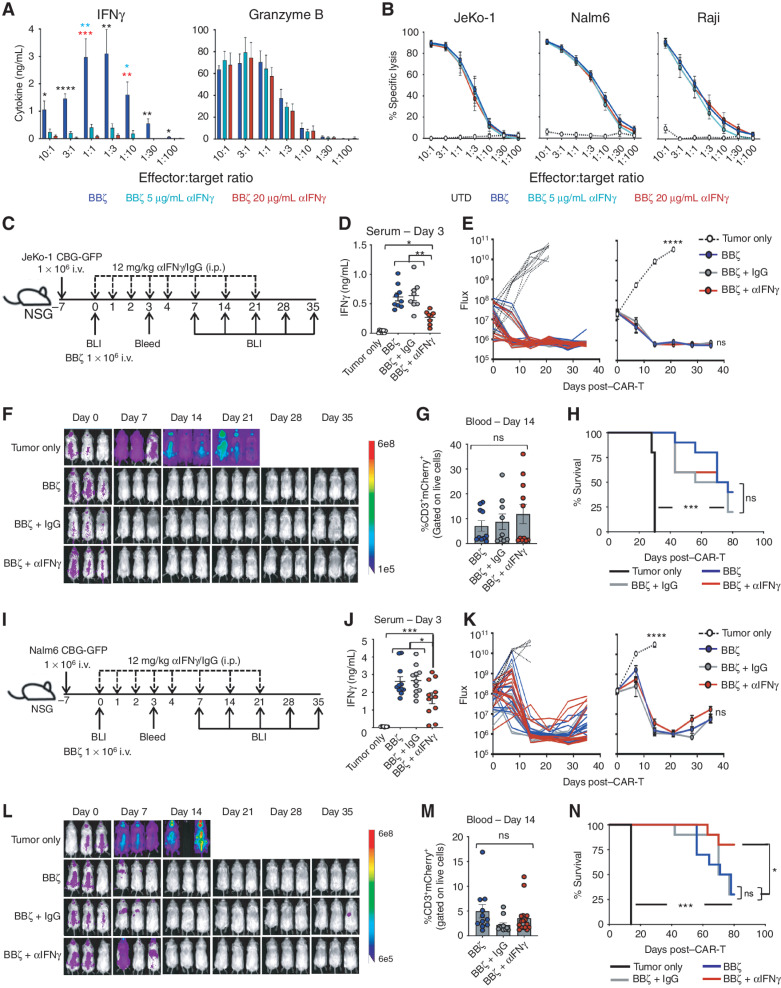
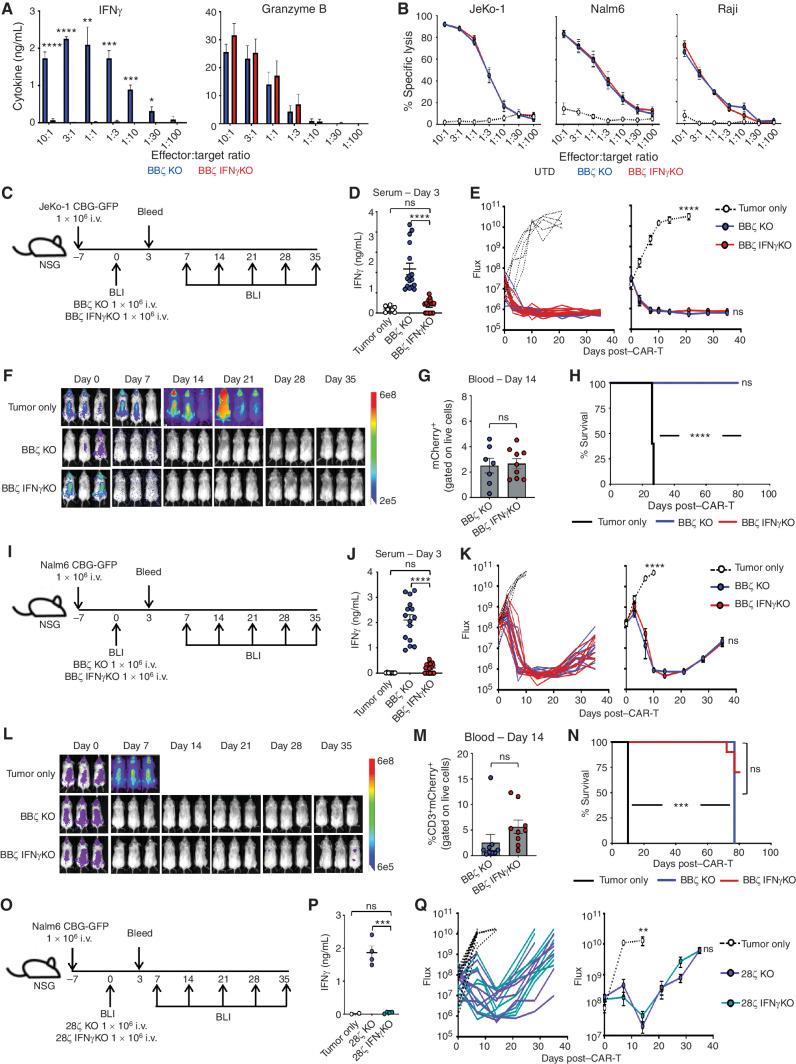
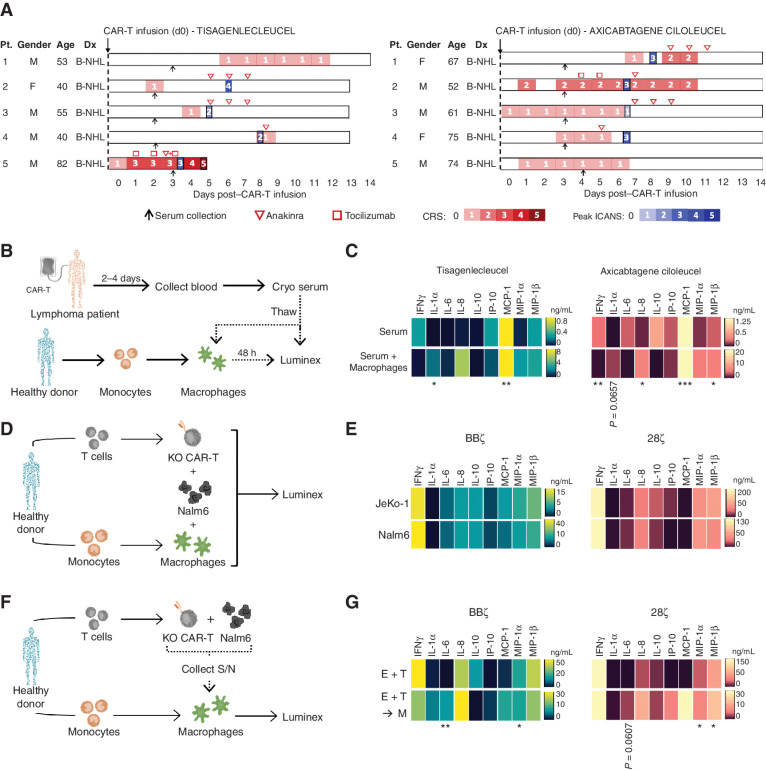
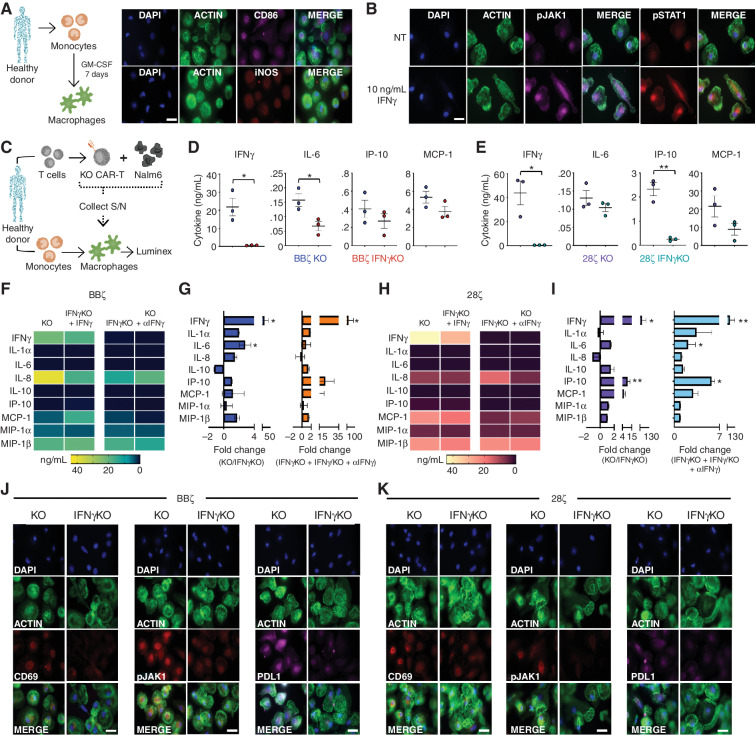
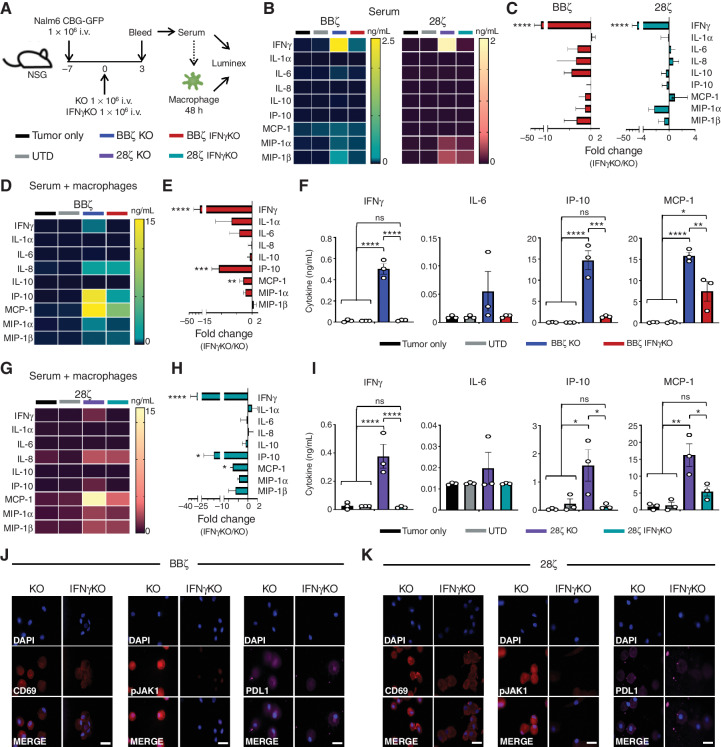
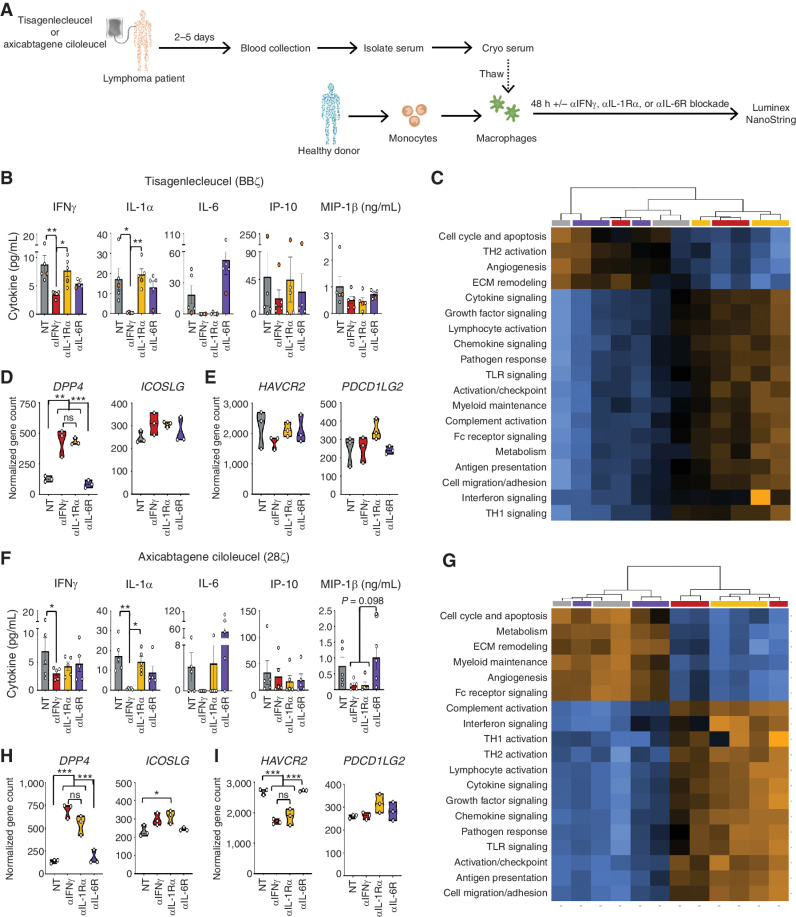
Chimeric antigen receptor (CAR) T cells induce impressive responses in patients with hematologic malignancies but can also trigger cytokine release syndrome (CRS), a systemic toxicity caused by activated CAR T cells and innate immune cells. Although IFNγ production serves as a potency assay for CAR T cells, its biologic role in conferring responses in hematologic malignancies is not established. Here we show that pharmacologic blockade or genetic knockout of IFNγ reduced immune checkpoint protein expression with no detrimental effect on antitumor efficacy against hematologic malignancies in vitro or in vivo. Furthermore, IFNγ blockade reduced macrophage activation to a greater extent than currently used cytokine antagonists in immune cells from healthy donors and serum from patients with CAR T-cell-treated lymphoma who developed CRS. Collectively, these data show that IFNγ is not required for CAR T-cell efficacy against hematologic malignancies, and blocking IFNγ could simultaneously mitigate cytokine-related toxicities while preserving persistence and antitumor efficacy.
Significance: Blocking IFNγ in CAR T cells does not impair their cytotoxicity against hematologic tumor cells and paradoxically enhances their proliferation and reduces macrophage-mediated cytokines and chemokines associated with CRS. These findings suggest that IFNγ blockade may improve CAR T-cell function while reducing treatment-related toxicity in hematologic malignancies. See related content by McNerney et al., p. 90 (17). This article is highlighted in the In This Issue feature, p. 85.
©2021 American Association for Cancer Research.
Figures
Comment in
-
Potential Role of IFNγ Inhibition in Refractory Cytokine Release Syndrome Associated with CAR T-cell Therapy.Blood Cancer Discov. 2022 Mar 1;3(2):90-94. doi: 10.1158/2643-3230.BCD-21-0203. Blood Cancer Discov. 2022. PMID: 35015687 Free PMC article.
Comment on
-
Potential Role of IFNγ Inhibition in Refractory Cytokine Release Syndrome Associated with CAR T-cell Therapy.Blood Cancer Discov. 2022 Mar 1;3(2):90-94. doi: 10.1158/2643-3230.BCD-21-0203. Blood Cancer Discov. 2022. PMID: 35015687 Free PMC article.
References
-
- Wu C, Xue Y, Wang P, Lin L, Liu Q, Li Net al. IFN-gamma primes macrophage activation by increasing phosphatase and tensin homolog via downregulation of miR-3473b. J Immunol 2014;193:3036–44. - PubMed
-
- Wang XB, Zheng CY, Giscombe R, Lefvert AK. Regulation of surface and intracellular expression of CTLA-4 on human peripheral T cells. Scand J Immunol 2001;54:453–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
