

The Mechanism of Action of Ginkgolic Acid (15:1) against Gram-Positive Bacteria Involves Cross Talk with Iron Homeostasis
- PMID: 35019708
- PMCID: PMC8754111
- DOI: 10.1128/spectrum.00991-21
The Mechanism of Action of Ginkgolic Acid (15:1) against Gram-Positive Bacteria Involves Cross Talk with Iron Homeostasis
Abstract
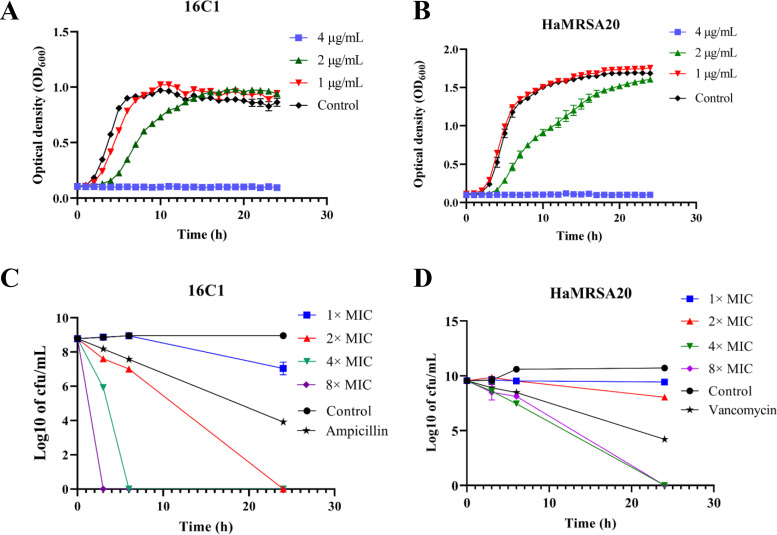
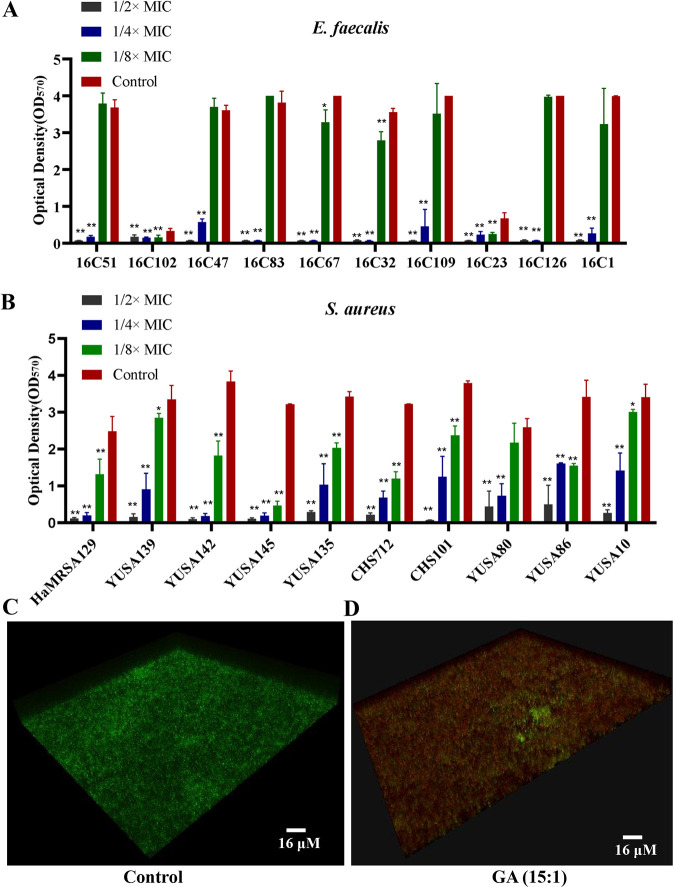
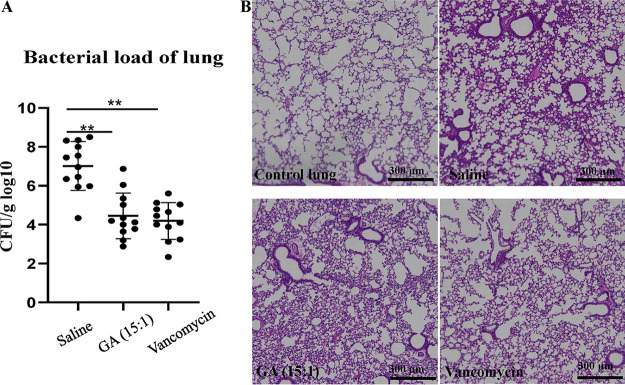
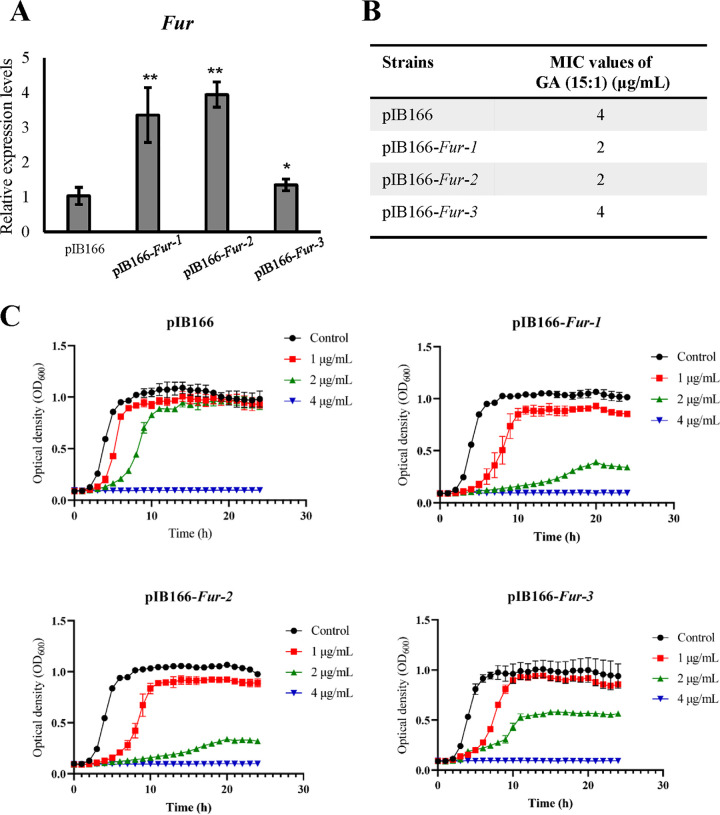
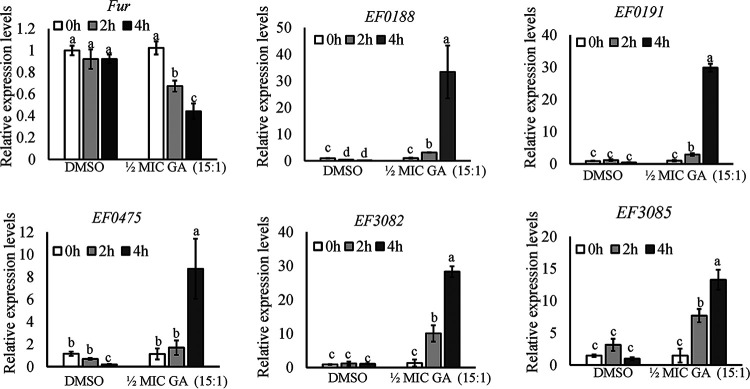
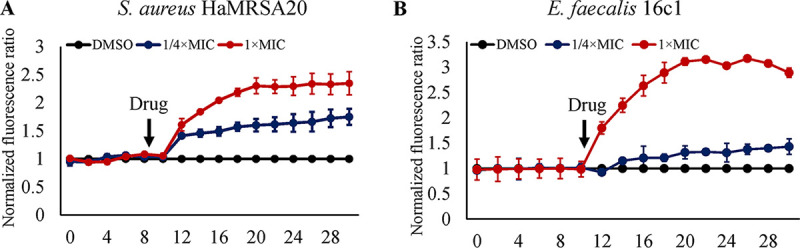
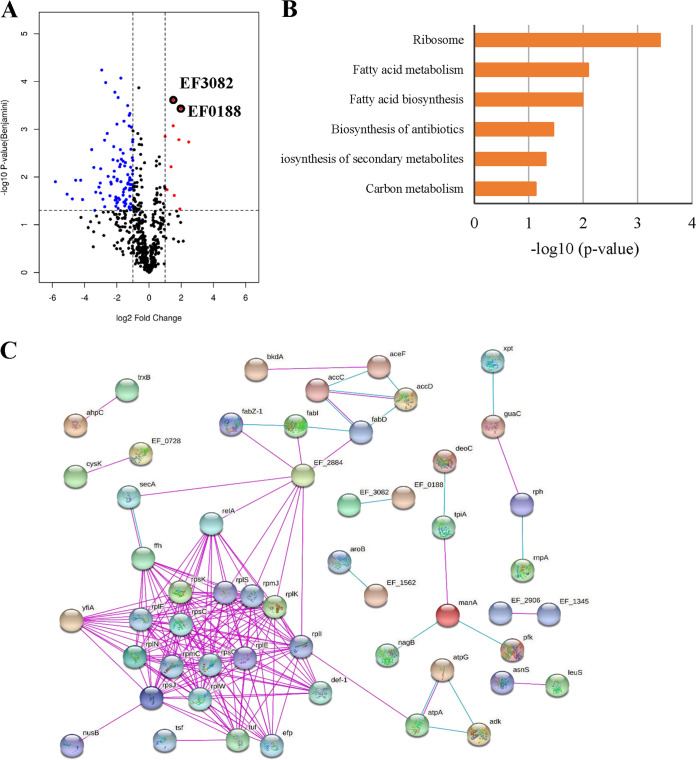
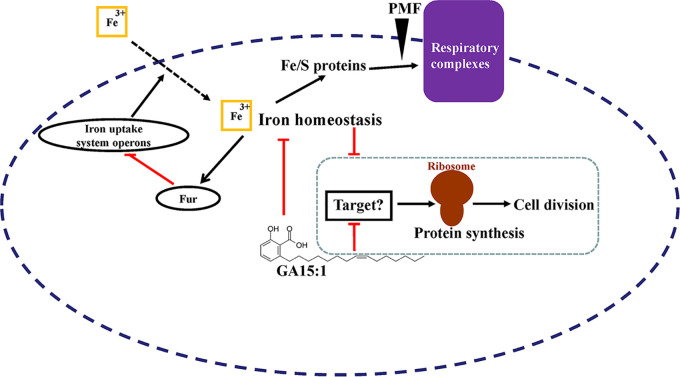
With the increasing reports of community-acquired and nosocomial infection caused by multidrug-resistant Gram-positive pathogens, there is an urgent need to develop new antimicrobial agents with novel antibacterial mechanisms. Here, we investigated the antibacterial activity of the natural product ginkgolic acid (GA) (15:1), derived from Ginkgo biloba, and its potential mode of action against the Gram-positive bacteria Enterococcus faecalis and Staphylococcus aureus. The MIC values of GA (15:1) against clinical E. faecalis and S. aureus isolates from China were ≤4 and ≤8 μg/mL, respectively, from our test results. Moreover, GA (15:1) displayed high efficiency in biofilm formation inhibition and bactericidal activity against E. faecalis and S. aureus. During its inhibition of the planktonic bacteria, the antibacterial activity of GA (15:1) was significantly improved under the condition of abolishing iron homeostasis. When iron homeostasis was abolished, inhibition of planktonic bacteria by GA (15:1) was significantly improved. This phenomenon can be interpreted as showing that iron homeostasis disruption facilitated the disruption of the functions of ribosome and protein synthesis by GA (15:1), resulting in inhibition of bacterial growth and cell death. Genetic mutation of ferric uptake regulator (Fur) led to GA (15:1) tolerance in in vitro-induced resistant derivatives, while overexpression of Fur led to increased GA (15:1) susceptibility. Additionally, GA (15:1) significantly decreased the bacterial loads of S. aureus strain USA300 in the lung tissues of mice in a pneumonic murine model. Conclusively, this study revealed an antimicrobial mechanism of GA (15:1) involving cross talk with iron homeostasis against Gram-positive pathogens. In the future, the natural product GA (15:1) might be applied to combat infections caused by Gram-positive pathogens. IMPORTANCE The increasing emergence of infectious diseases associated with multidrug-resistant Gram-positive pathogens has raised the urgent need to develop novel antibiotics. GA (15:1) is a natural product derived from Ginkgo biloba and possesses a wide range of bioactivities, including antimicrobial activity. However, its antibacterial mechanisms remain unclear. Our current study found that the function of ferric uptake regulator (Fur) was highly correlated with the antimicrobial activity of GA (15:1) against E. faecalis and that the antibacterial activity of GA (15:1) could be strengthened by the disruption of iron homeostasis. This study provided important insight into the mode of action of GA (15:1) against Gram-positive bacteria and suggested that GA (15:1) holds the potential to be an antimicrobial treatment option for infection caused by multidrug-resistant Gram-positive pathogens.
Keywords: E. faecalis; Fur; antibacterial mechanism; ferric uptake regulator; ginkgolic acid (15:1); iron homeostasis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Inagaki K, Lucar J, Blackshear C, Hobbs CV. 2019. Methicillin-susceptible and methicillin-resistant Staphylococcus aureus bacteremia: nationwide estimates of 30-day readmission, in-hospital mortality, length of stay, and cost in the United States. Clin Infect Dis 69:2112–2118. doi:10.1093/cid/ciz123. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
