

Multiomic Profiling of Central Nervous System Leukemia Identifies mRNA Translation as a Therapeutic Target
- PMID: 35019858
- PMCID: PMC9783958
- DOI: 10.1158/2643-3230.BCD-20-0216
Multiomic Profiling of Central Nervous System Leukemia Identifies mRNA Translation as a Therapeutic Target
Abstract
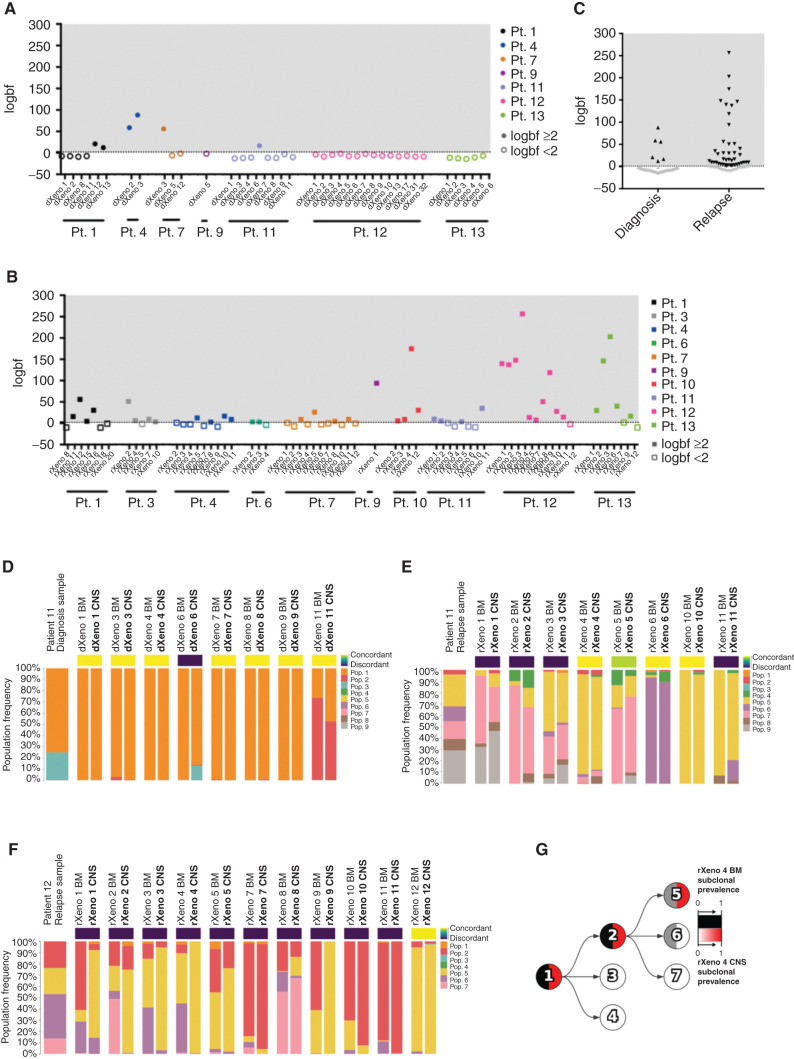
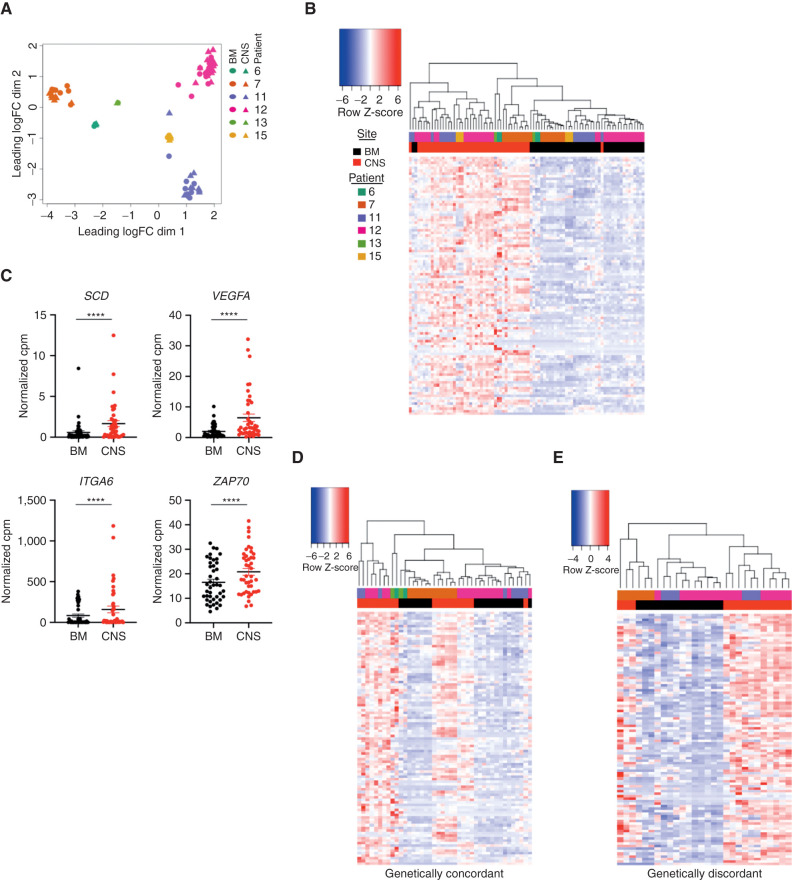
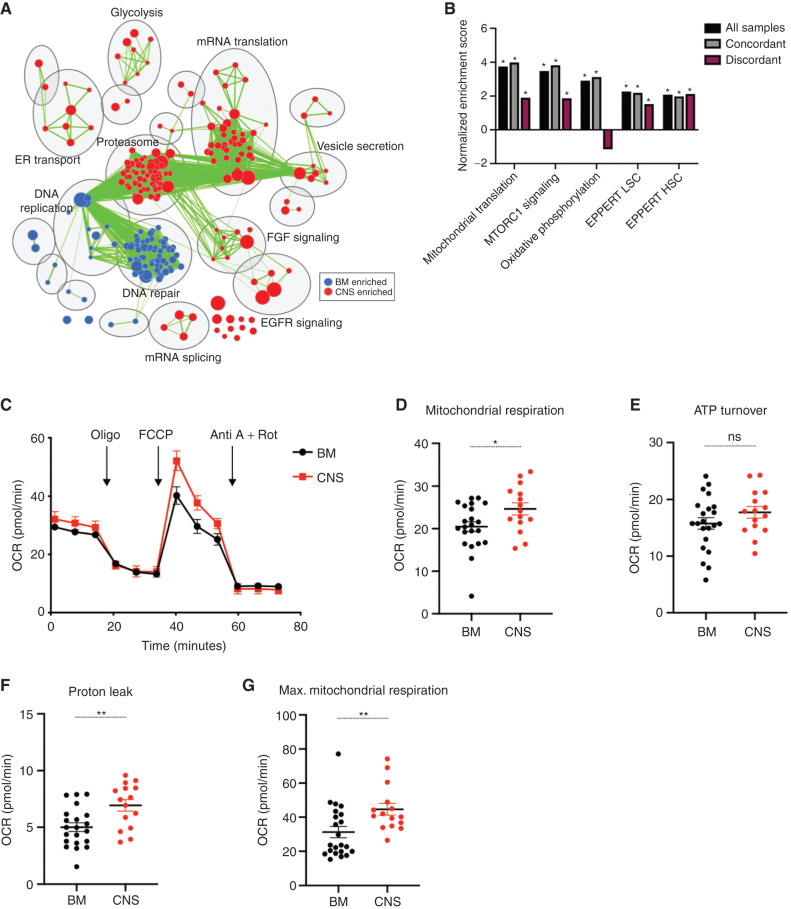
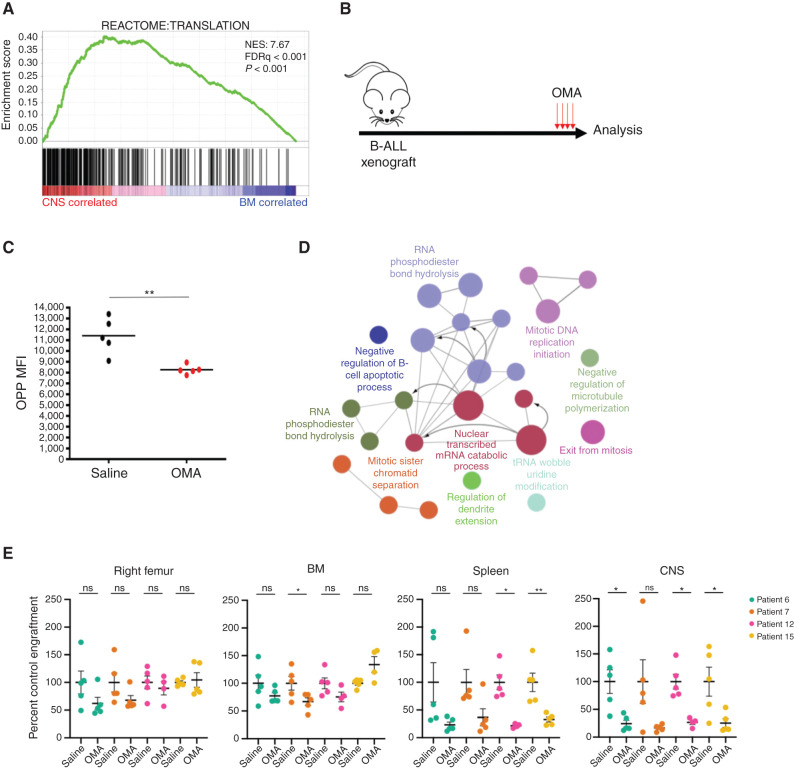
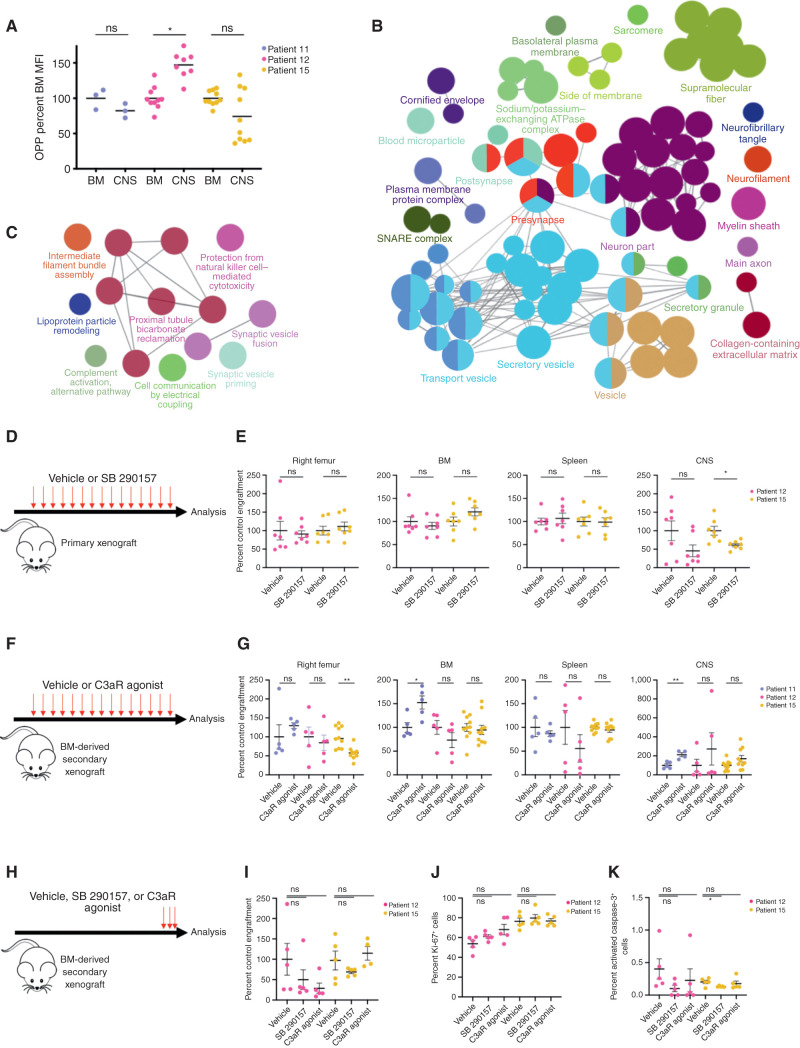
Central nervous system (CNS) dissemination of B-precursor acute lymphoblastic leukemia (B-ALL) has poor prognosis and remains a therapeutic challenge. Here we performed targeted DNA sequencing as well as transcriptional and proteomic profiling of paired leukemia-infiltrating cells in the bone marrow (BM) and CNS of xenografts. Genes governing mRNA translation were upregulated in CNS leukemia, and subclonal genetic profiling confirmed this in both BM-concordant and BM-discordant CNS mutational populations. CNS leukemia cells were exquisitely sensitive to the translation inhibitor omacetaxine mepesuccinate, which reduced xenograft leptomeningeal disease burden. Proteomics demonstrated greater abundance of secreted proteins in CNS-infiltrating cells, including complement component 3 (C3), and drug targeting of C3 influenced CNS disease in xenografts. CNS-infiltrating cells also exhibited selection for stemness traits and metabolic reprogramming. Overall, our study identifies targeting of mRNA translation as a potential therapeutic approach for B-ALL leptomeningeal disease. SIGNIFICANCE: Cancer metastases are often driven by distinct subclones with unique biological properties. Here we show that in B-ALL CNS disease, the leptomeningeal environment selects for cells with unique functional dependencies. Pharmacologic inhibition of mRNA translation signaling treats CNS disease and offers a new therapeutic approach for this condition.This article is highlighted in the In This Issue feature, p. 1.
©2021 American Association for Cancer Research.
Figures
Comment in
- 2643-3230. doi: 10.1158/2643-3230.BCD-3-1-ITI doi: 10.1158/2643-3230.BCD-3-1-ITI
References
-
- Gaudichon J, Jakobczyk H, Debaize L, Cousin E, Galibert MD, Troadec MBet al. Mechanisms of extramedullary relapse in acute lymphoblastic leukemia: reconciling biological concepts and clinical issues. Blood Rev 2019;36:40–56. - PubMed
-
- Pui CH, Howard SC. Current management and challenges of malignant disease in the CNS in paediatric leukaemia. Lancet Oncol 2008;9:257–68. - PubMed
-
- Fielding AK, Richards SM, Chopra R, Lazarus HM, Litzow MR, Buck Get al. Outcome of 609 adults after relapse of acute lymphoblastic leukemia (ALL); an MRC UKALL12/ECOG 2993 study. Blood 2007;109:944–50. - PubMed
-
- Hunger SP, Mullighan CG. Acute lymphoblastic leukemia in children. N Engl J Med 2015;373:1541–52. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
