

Smooth muscle mineralocorticoid receptor as an epigenetic regulator of vascular ageing
- PMID: 35020830
- PMCID: PMC10060709
- DOI: 10.1093/cvr/cvac007
Smooth muscle mineralocorticoid receptor as an epigenetic regulator of vascular ageing
Abstract
Aims: Vascular stiffness increases with age and independently predicts cardiovascular disease risk. Epigenetic changes, including histone modifications, accumulate with age but the global pattern has not been elucidated nor are the regulators known. Smooth muscle cell-mineralocorticoid receptor (SMC-MR) contributes to vascular stiffness in ageing mice. Thus, we investigated the regulatory role of SMC-MR in vascular epigenetics and stiffness.
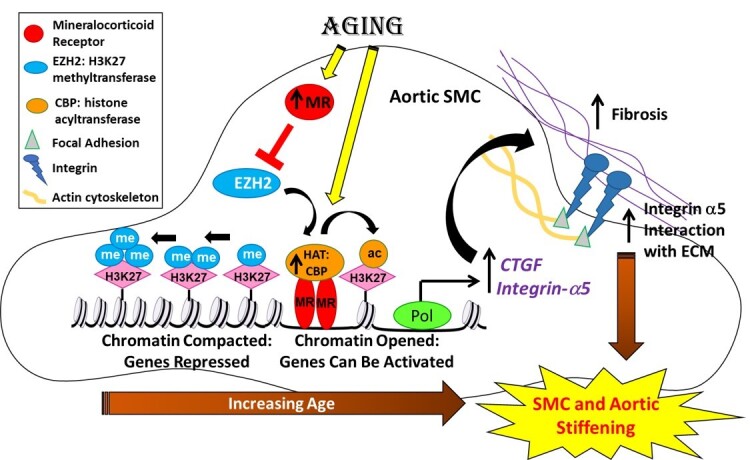
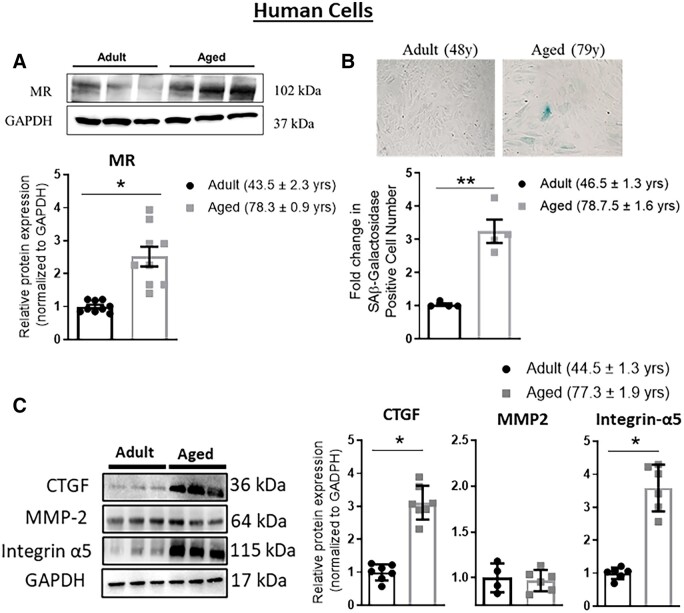
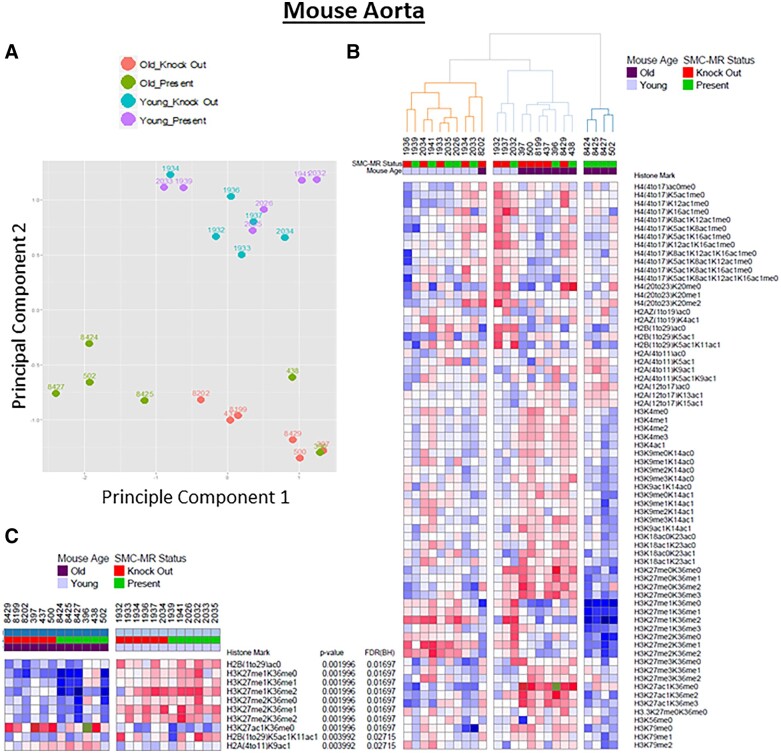
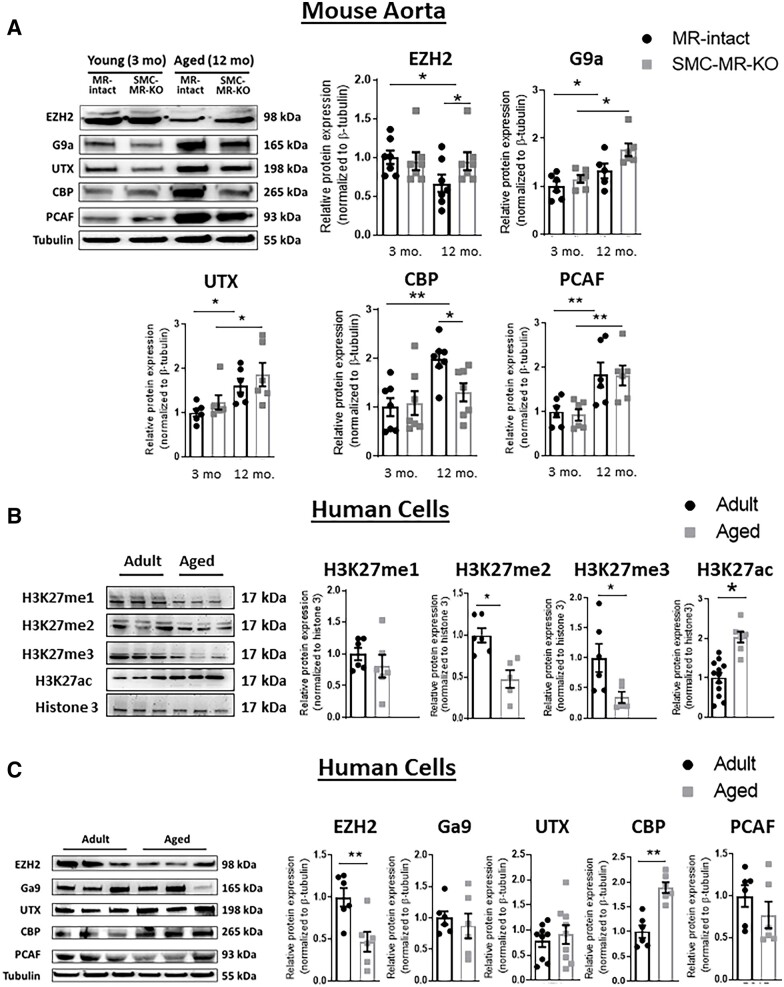
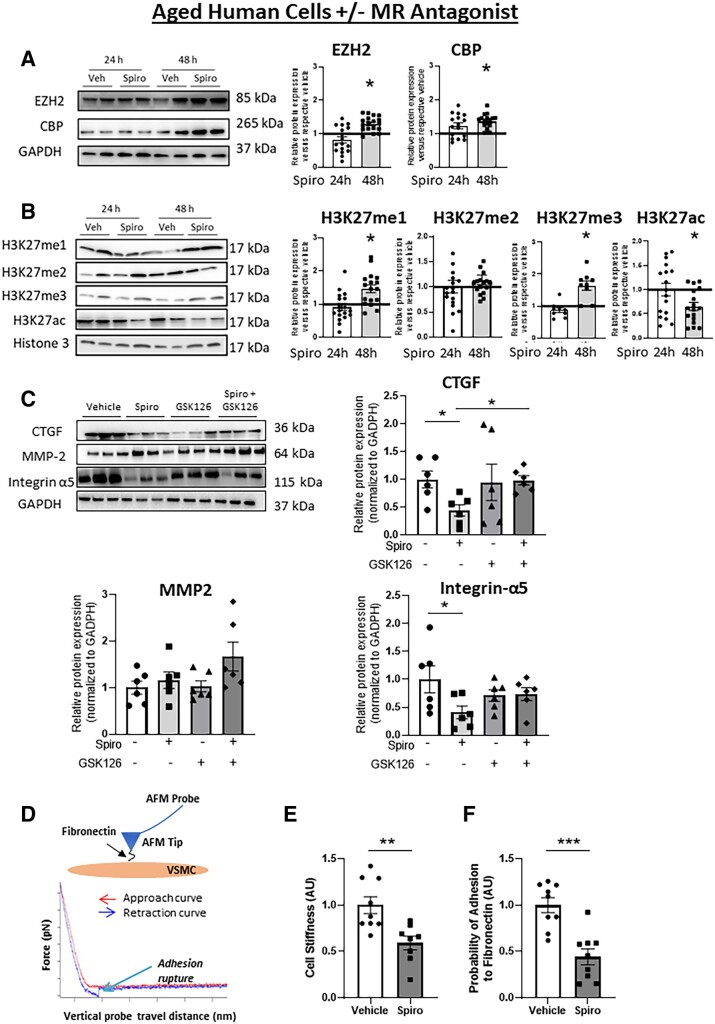
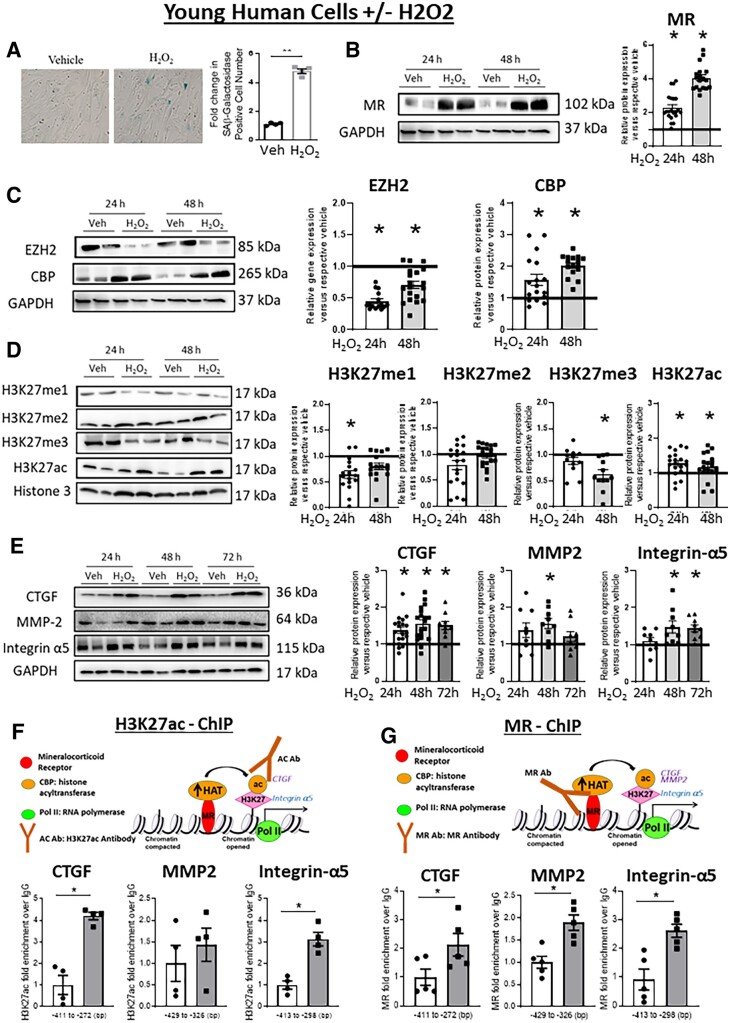
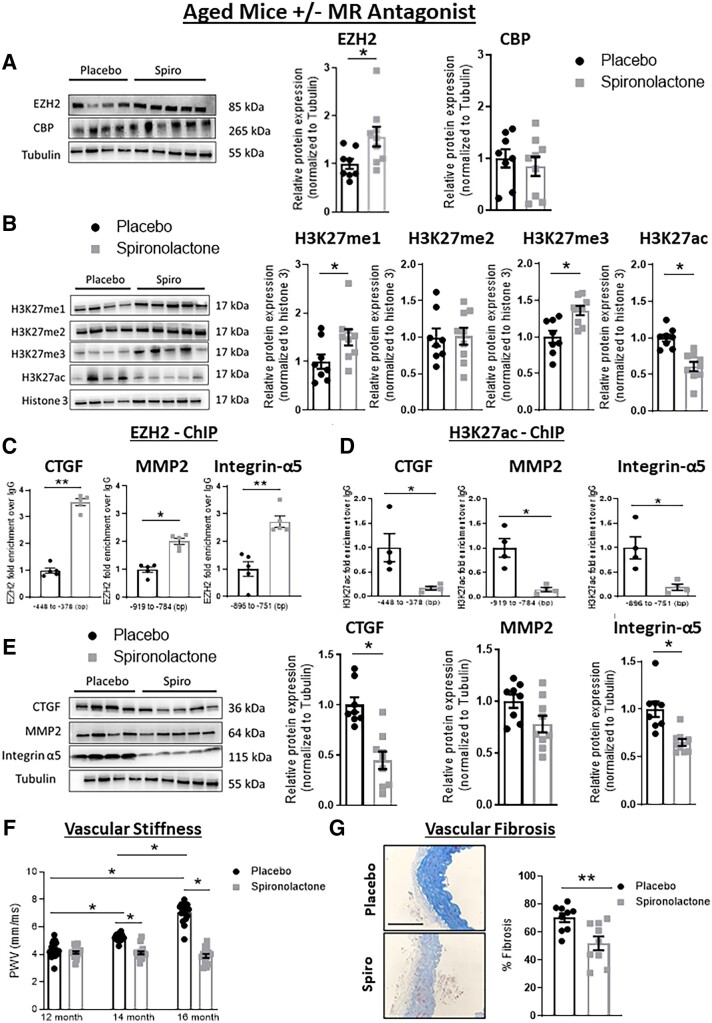
Methods and results: Mass spectrometry-based proteomic profiling of all histone modifications completely distinguished 3 from 12-month-old mouse aortas. Histone-H3 lysine-27 (H3K27) methylation (me) significantly decreased in ageing vessels and this was attenuated in SMC-MR-KO littermates. Immunoblotting revealed less H3K27-specific methyltransferase EZH2 with age in MR-intact but not SMC-MR-KO vessels. These ageing changes were examined in primary human aortic (HA)SMC from adult vs. aged donors. MR, H3K27 acetylation (ac), and stiffness gene (connective tissue growth factor, integrin-α5) expression significantly increased, while H3K27me and EZH2 decreased, with age. MR inhibition reversed these ageing changes in HASMC and the decline in stiffness genes was prevented by EZH2 blockade. Atomic force microscopy revealed that MR antagonism decreased intrinsic stiffness and the probability of fibronectin adhesion of aged HASMC. Conversely, ageing induction in young HASMC with H2O2; increased MR, decreased EZH2, enriched H3K27ac and MR at stiffness gene promoters by chromatin immunoprecipitation, and increased stiffness gene expression. In 12-month-old mice, MR antagonism increased aortic EZH2 and H3K27 methylation, increased EZH2 recruitment and decreased H3K27ac at stiffness genes promoters, and prevented ageing-induced vascular stiffness and fibrosis. Finally, in human aortic tissue, age positively correlated with MR and stiffness gene expression and negatively correlated with H3K27me3 while MR and EZH2 are negatively correlated.
Conclusion: These data support a novel vascular ageing model with rising MR in human SMC suppressing EZH2 expression thereby decreasing H3K27me, promoting MR recruitment and H3K27ac at stiffness gene promoters to induce vascular stiffness and suggests new targets for ameliorating ageing-associated vascular disease.
Keywords: EZH2; Epigenetics; Vascular ageing; Vascular stiffness; Mineralocorticoid receptor.
Published on behalf of the European Society of Cardiology. All rights reserved. © The Author(s) 2022. For permissions, please email: journals.permissions@oup.com.
Conflict of interest statement
Conflict of interest: none declared. JDJ is currently employed by Odyssey Therapeutics. His contribution to the manuscript was completed while he was at the Broad Institute. There is no overlap between the subject of the manuscript and the role of JDJ at Odyssey Therapuetics.
Figures

References
-
- Mitchell GF, Parise H, Benjamin EJ, Larson MG, Keyes MJ, Vita JA, Vasan RS, Levy D. Changes in arterial stiffness and wave reflection with advancing age in healthy men and women: the Framingham Heart Study. Hypertension 2004;43:1239–1245. - PubMed
-
- Cecelja M, Shanahan CM. Targeting cell stiffness: a paradigm shift in the treatment of aortic stiffness. Circ Res 2021;128:769–771. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
