

ATRX loss in glioma results in dysregulation of cell-cycle phase transition and ATM inhibitor radio-sensitization
- PMID: 35021084
- PMCID: PMC8759735
- DOI: 10.1016/j.celrep.2021.110216
ATRX loss in glioma results in dysregulation of cell-cycle phase transition and ATM inhibitor radio-sensitization
Abstract
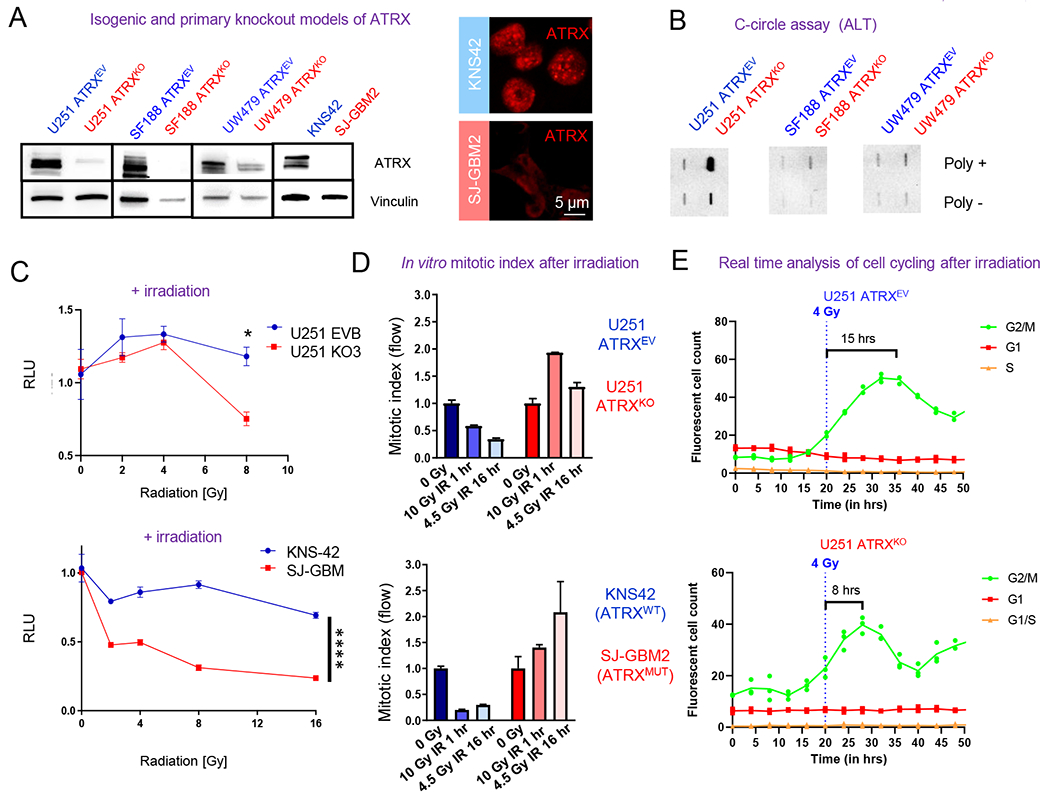
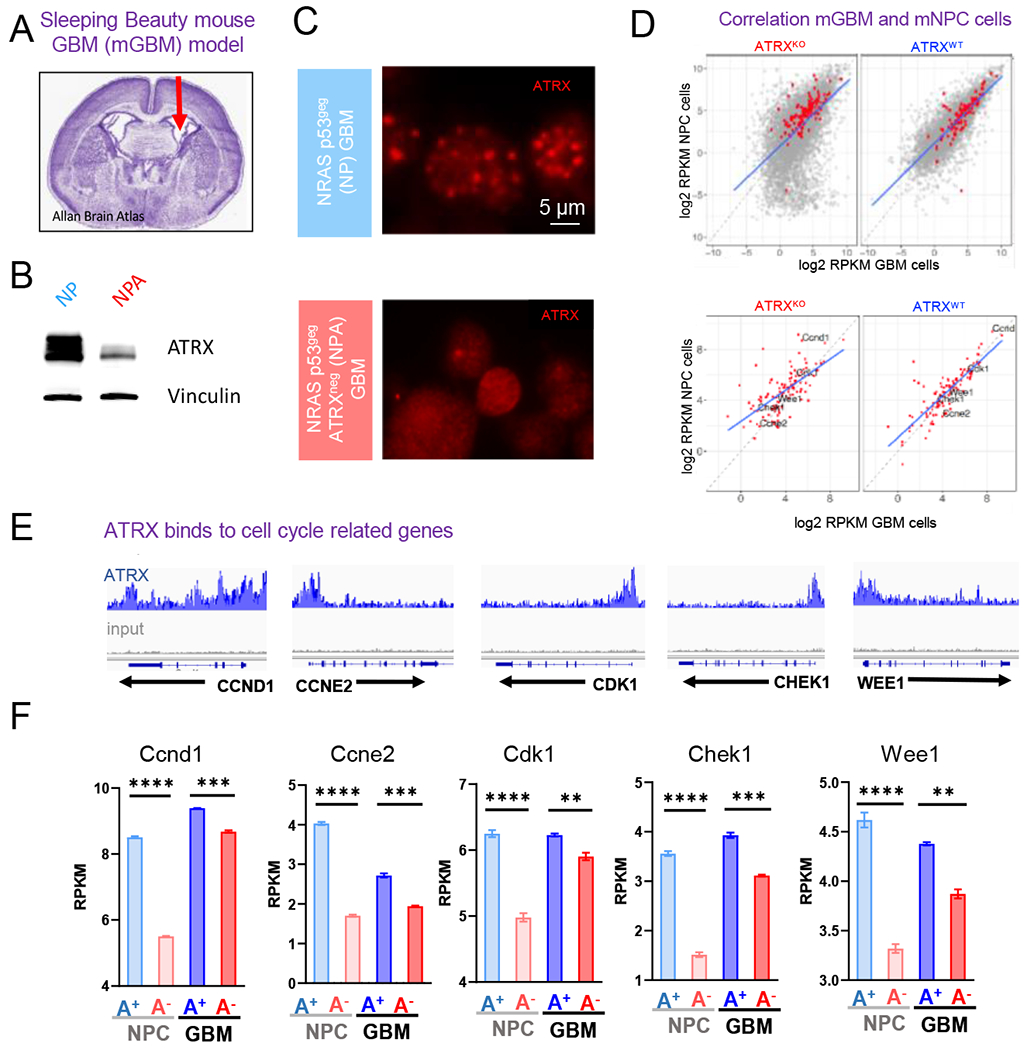
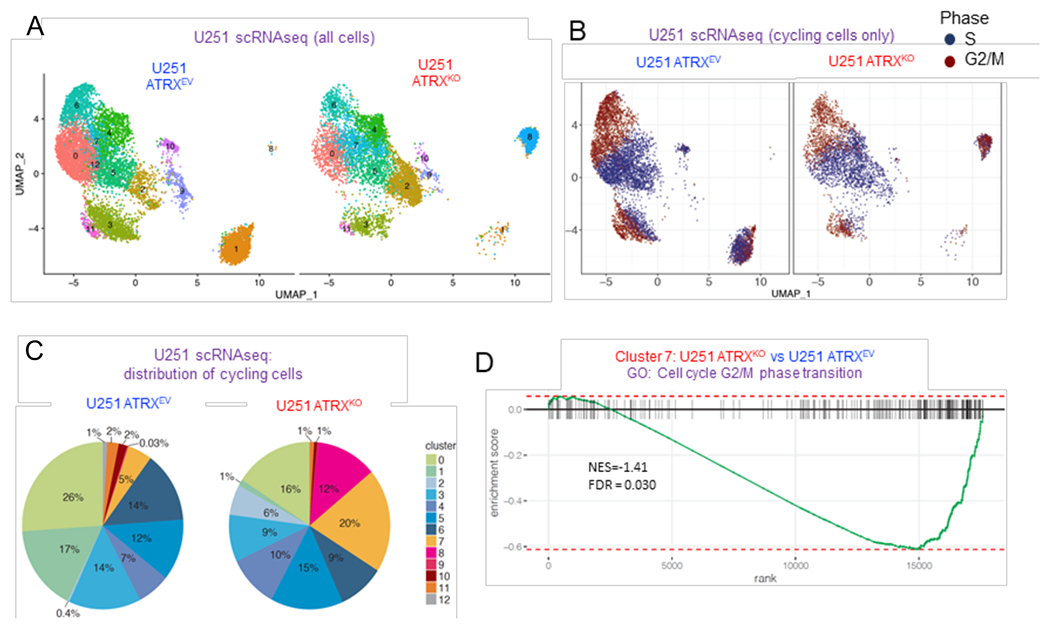
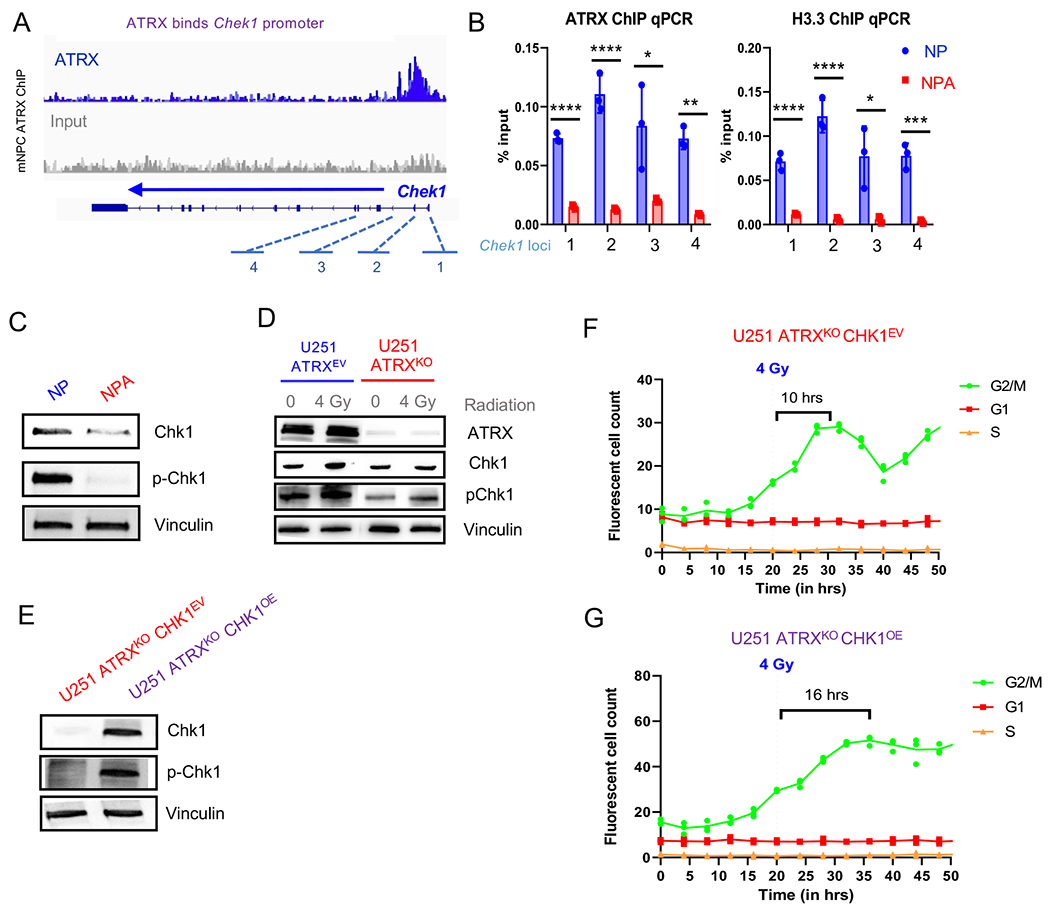
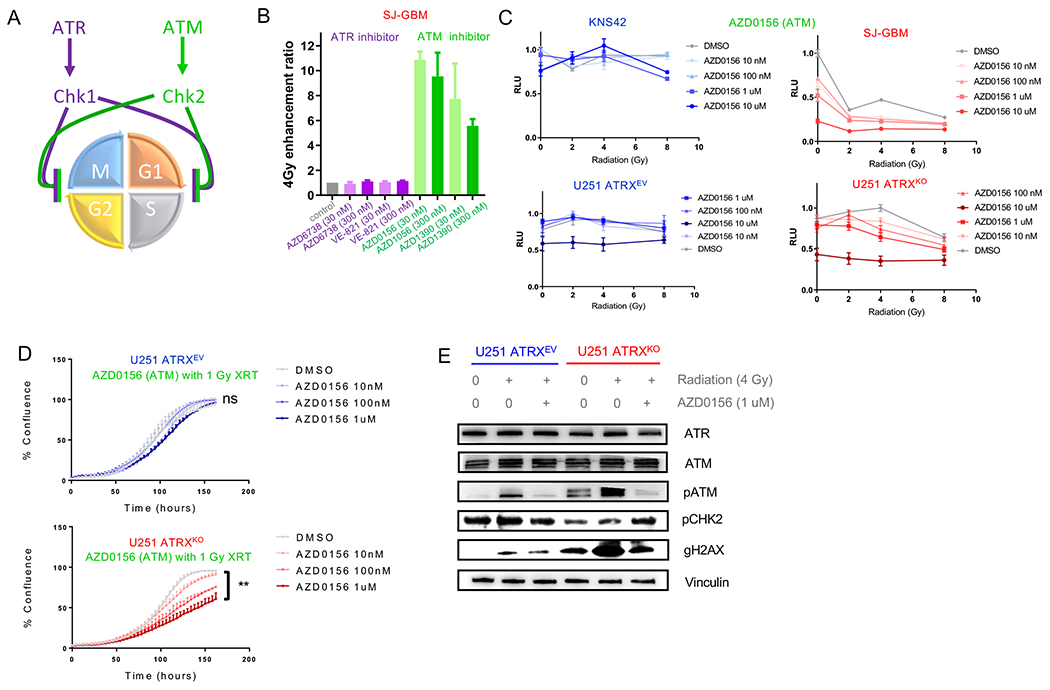
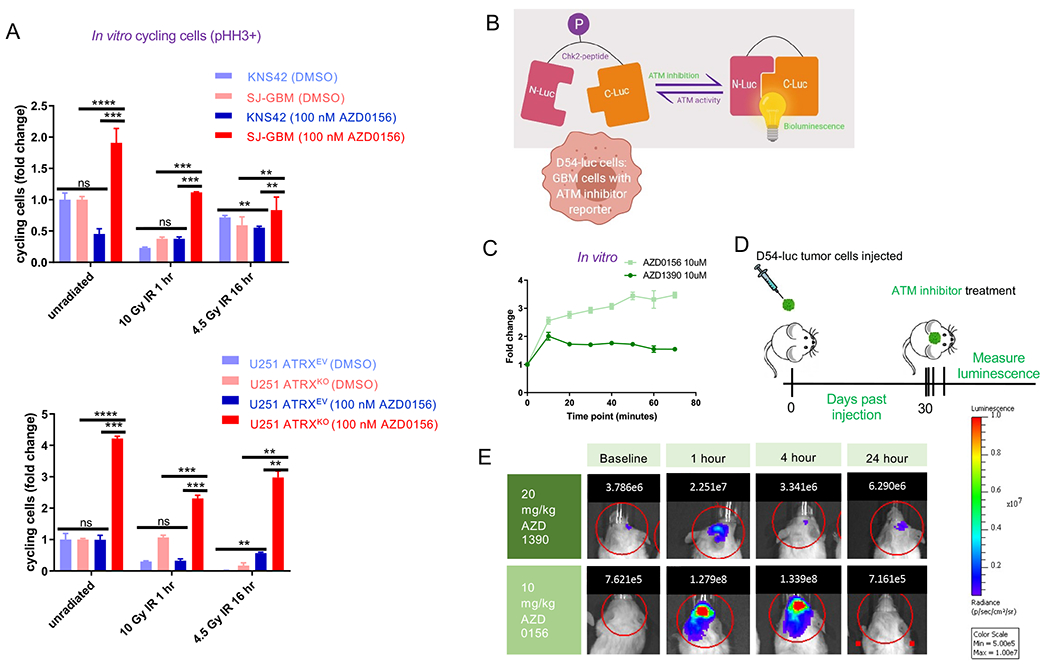
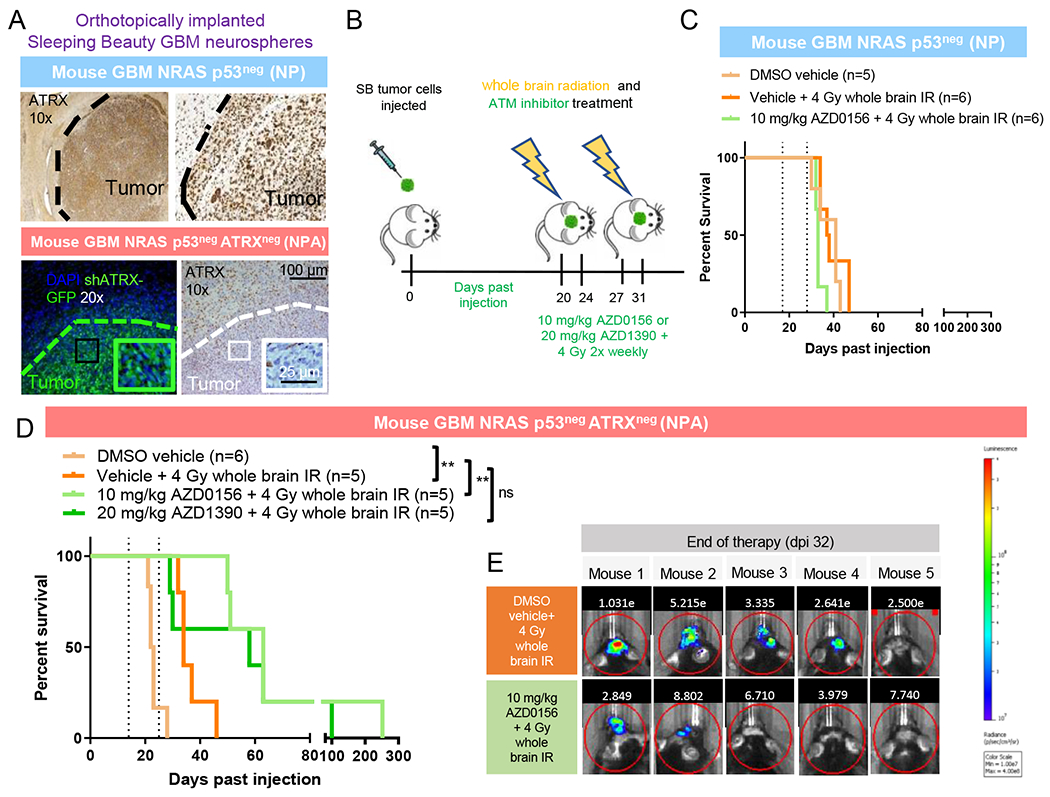
ATRX, a chromatin remodeler protein, is recurrently mutated in H3F3A-mutant pediatric glioblastoma (GBM) and isocitrate dehydrogenase (IDH)-mutant grade 2/3 adult glioma. Previous work has shown that ATRX-deficient GBM cells show enhanced sensitivity to irradiation, but the etiology remains unclear. We find that ATRX binds the regulatory elements of cell-cycle phase transition genes in GBM cells, and there is a marked reduction in Checkpoint Kinase 1 (CHEK1) expression with ATRX loss, leading to the early release of G2/M entry after irradiation. ATRX-deficient cells exhibit enhanced activation of master cell-cycle regulator ATM with irradiation. Addition of the ATM inhibitor AZD0156 doubles median survival in mice intracranially implanted with ATRX-deficient GBM cells, which is not seen in ATRX-wild-type controls. This study demonstrates that ATRX-deficient high-grade gliomas (HGGs) display Chk1-mediated dysregulation of cell-cycle phase transitions, which opens a window for therapies targeting this phenotype.
Keywords: ATM inhibitor; ATRX; CHEK1; cell-cycle; epigenetics; glioma.
Copyright © 2021. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Andersson R, Gebhard C, Miguel-Escalada I, Hoof I, Bornholdt J, Boyd M, Chen Y, Zhao X, Andrews S (2010). FastQC: a quality control tool for high throughput sequence data. Version 0.11. 2 Website: http://www.bioinformatics.babraham.ac.uk/projects/fastqc.
-
- Bender S, Tang Y, Lindroth AM, Hovestadt V, Jones DT, Kool M, Zapatka M, Northcott PA, Sturm D, Wang W, et al. (2013). Reduced H3K27me3 and DNA hypomethylation are major drivers of gene expression in K27M mutant pediatric high-grade gliomas. Cancer Cell 24, 660–672. 10.1016/j.ccr.2013.10.006. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS122165/NS/NINDS NIH HHS/United States
- R01 NS105556/NS/NINDS NIH HHS/United States
- U01 CA216449/CA/NCI NIH HHS/United States
- R01 NS082311/NS/NINDS NIH HHS/United States
- P30 CA046592/CA/NCI NIH HHS/United States
- R01 NS110572/NS/NINDS NIH HHS/United States
- R01 CA240515/CA/NCI NIH HHS/United States
- T32 CA009676/CA/NCI NIH HHS/United States
- P30 ES017885/ES/NIEHS NIH HHS/United States
- R37 NS094804/NS/NINDS NIH HHS/United States
- R01 NS122234/NS/NINDS NIH HHS/United States
- K08 NS099427/NS/NINDS NIH HHS/United States
- R21 NS107894/NS/NINDS NIH HHS/United States
- R01 NS119231/NS/NINDS NIH HHS/United States
- R01 NS096756/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
