

LRRTM3 regulates activity-dependent synchronization of synapse properties in topographically connected hippocampal neural circuits
- PMID: 35022233
- PMCID: PMC8784129
- DOI: 10.1073/pnas.2110196119
LRRTM3 regulates activity-dependent synchronization of synapse properties in topographically connected hippocampal neural circuits
Abstract
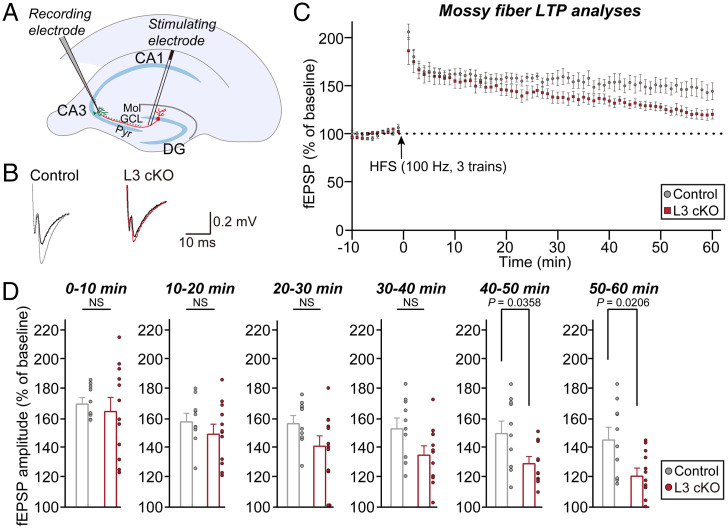
Synaptic cell-adhesion molecules (CAMs) organize the architecture and properties of neural circuits. However, whether synaptic CAMs are involved in activity-dependent remodeling of specific neural circuits is incompletely understood. Leucine-rich repeat transmembrane protein 3 (LRRTM3) is required for the excitatory synapse development of hippocampal dentate gyrus (DG) granule neurons. Here, we report that Lrrtm3-deficient mice exhibit selective reductions in excitatory synapse density and synaptic strength in projections involving the medial entorhinal cortex (MEC) and DG granule neurons, accompanied by increased neurotransmitter release and decreased excitability of granule neurons. LRRTM3 deletion significantly reduced excitatory synaptic innervation of hippocampal mossy fibers (Mf) of DG granule neurons onto thorny excrescences in hippocampal CA3 neurons. Moreover, LRRTM3 loss in DG neurons significantly decreased mossy fiber long-term potentiation (Mf-LTP). Remarkably, silencing MEC-DG circuits protected against the decrease in the excitatory synaptic inputs onto DG and CA3 neurons, excitability of DG granule neurons, and Mf-LTP in Lrrtm3-deficient mice. These results suggest that LRRTM3 may be a critical factor in activity-dependent synchronization of the topography of MEC-DG-CA3 excitatory synaptic connections. Collectively, our data propose that LRRTM3 shapes the target-specific structural and functional properties of specific hippocampal circuits.
Keywords: LRRTM3; dentate gyrus; excitatory synapse; long-term plasticity; medial entorhinal cortex.
Copyright © 2022 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
